Skala czasu geologicznego
Era : Okres : Epoka : Miliony lat temu (w przybliżeniu) : Najważniejsze wydarzenia
Kenozoik : czwartorzęd : holocen : 0,01 : koniec ostatniej epoki lodowcowej i początki rolnictwa, udomowienia zwierząt i cywilizacji na dużą skalę
: : Plejstocen : 1.8 : Epoki lodowcowe . Rozproszenie homininów z Afryki
: Neogene : pliocen : 5 : Pojawienie się dwunożnych homininów. Małpy promieniują. Pojawiają się duże drapieżniki
: : miocen: małpy ewoluują i promieniują
: Paleogen : oligocen : 35 : małpy i duże biegające ssaki wyłaniają się
: : eocen : 54 : Promieniowanie współczesnych typów ssaków
: : paleocen : 65 : Pierwsze ssaki łożyskowe. Pierwsze naczelne
Mezozoik : kreda : 145 : Pierwsze rośliny kwitnące. Dinozaury osiągają szczyt, a następnie wymierają pod koniec kredy.
: trias : 250 : Pierwsze dinozaury. Drzewa iglaste obfite
Paleozoik : perm : 290 : Pod koniec . Trylobity permskie giną podczas ogromnego wymierania
: karbon : 350 : Pierwsze gady. Imponujące lasy węglowe
: dewon : 400 : rekiny i płazy dominują i występują licznie
: sylur : 440 Pierwsze rośliny i zwierzęta lądowe
: ordowik : 500 : Dominują bezkręgowce. Pierwsze ryby
: kambr : 540 : "eksplozja" życia zwierzęcego w morzach, w tym trylobity
Prekambr : : 4600 : Życie na Ziemi się zaczyna. Rzadkie skamieniałości prymitywnych roślin wodnych. Niektóre algi kopalne mają co najmniej 2,5 miliarda lat
Początki Człowieków (67)
PRZEWIJANIE I POWTARZANIE EWOLUCJI
Co nam pozostało do myślenia o ludzkiej egzystencji? Ze wszystkimi dowodami wskazującymi na nasze małpopodobne pochodzenie 6 Mya i nasze wspólne pochodzenie ze wszystkim, począwszy od karaluchów, śluzowców, szumowin stawowych, ptaszników, żmij żmijowych, grzybów stóp, a nawet wirusów, wyjątkowy status ewolucyjny H. sapiens może wydają się ponure, a może nawet obrzydliwe. Ale nawet jeśli mamy kiepskich krewnych, czy powinniśmy mniej myśleć o naszym gatunku? Jeśli zapomnimy o wspomnianych wcześniej negatywnych skutkach, jakie wywarliśmy na planecie, czy nadal można rozkoszować się naszą wyjątkową wyjątkowością? W końcu niezwykłe jest to, co Matka Natura stworzyła z przodka z małym mózgiem, niezdarnymi rękami i bez znaczących emocjonalnych lub artystycznych form ekspresji. Oboje jesteśmy dziwni i wspaniali, ale czy jesteśmy wyjątkowi na zawsze i na całą wieczność? Co by było, gdyby tylko jedno wydarzenie wydarzyło się inaczej w historii Ziemi - czy ludzie nadal ewoluowali? Gdyby asteroida nigdy nie zniszczyła dinozaurów, możliwe, że nie byłoby nas tutaj dzisiaj. Światowe wymieranie na granicy kredy i trzeciorzędu oszczędziło małych ssaków i to z tych ocalałych narodziło się całe promieniowanie ssaków. Dzięki tym wczesnym ssakom to żyrafy, a nie brontozaury, zjadają liście z najwyższych drzew i to ludzie, lwy i hieny, a nie tyranozaury, siedzą na szczycie piramidy troficznej. Rozważ ustawienie zegara z powrotem na zero, z powrotem do Wielkiego Wybuchu i powstania tego wszechświata 14 miliardów lat temu. Następnie wpraw go w ruch. Jak wyglądałoby "przeróbka" historii? Czy będzie to to samo, jak powtórka telewizyjna, czy też stanie się zupełnie innym odcinkiem? Paleontolodzy Simon Conway Morris i Stephen Jay Gould (1941-2002) weszli w słynną debatę na ten temat. Simon Conway Morris twierdzi, że powtórzenie ewolucji byłoby powtórką, ponieważ historia jest ograniczona prawami fizyki, biologii i tak dalej; nie wszystko jest możliwe. W normalnych warunkach środowiskowych życie odpowiednio się dostosuje i jest tylko tak wiele sposobów, aby to zrobić. Morris podkreśla liczne przykłady zbieżnej ewolucji, takie jak rekiny i delfiny, i ewolucja kotów szablozębnych zarówno w rodach torbaczy, jak i ssaków łożyskowych. Stephen Jay Gould widział całkowicie odwrotny pogląd i argumentował, że powtórka ewolucji zaowocuje zupełnie nowym epizodem życia. Nazwał ludzi i całą gildię żywych organizmów żyjących dzisiaj "chwalebnym wypadkiem" spowodowanym przez niepowtarzalną, niepowtarzalną sekwencję wydarzeń w historii Ziemi. Za każdym razem, gdy ewolucja jest przewijana i odtwarzana na nowo, a loteria wydarzeń zaczyna się od nowa, wynikiem będzie zupełnie nowy wynik z całkowicie unikalnymi organizmami. Chociaż przyznaje, że istnieją granice różnorodności, które mogą wyniknąć z takiego hipotetycznego eksperymentu, Gould twierdził, że szanse są prawie niemożliwe że dzisiejszy świat z dzisiejszymi stworzeniami nieuchronnie ewoluowałby, gdyby Matka Natura otrzymała kolejny wir. Według Goulda przykłady zbieżności są rzadkie i rozczarowujące, a najważniejsza cecha, o którą chodzi w jego argumentacji, wyjątkowość człowieczeństwa, człowieczeństwa i ludzkiej świadomości, nie rozwinęła się w żadnej innej żywej istocie. Z perspektywy Goulda trudno sobie wyobrazić, by coś takiego jak H. sapiens mogło kiedykolwiek ewoluować
Początki Człowieków (66)
GŁĘBOKI WPŁYW
Być może wyjątkowo ludzką cechą, która ma największy wpływ na świat, jest nasza skłonność do zmiany naszego środowiska. Jesteśmy ekspertami w tworzeniu śmieci i zostawiamy je wszędzie, na hałdach i kopcach. Nie możemy także podróżować daleko bez naszych stad bydła, owiec i kóz, które na naszej planecie liczą ponad 3 miliardy. Oramy pola, burzmy lasy i rozświetlamy nocne niebo. Gdziekolwiek pójdziemy, zmienia się rodzima flora i fauna. Niektóre czerpią zyski z naszego przybycia, jak szczury i ptaki, które żywią się naszymi śmieciami, ale inne kurczą się, a czasem znikają na dobre. Ludzie są tak zdolni do manipulowania środowiskiem, że zmieniliśmy nawet globalny klimat. Z pewnością globalne ocieplenie jest naturalnym procesem powodowanym przez cykle lodowcowe, ale spalanie paliw kopalnych emituje do atmosfery zanieczyszczenia, takie jak dwutlenek węgla, gdzie zostaje uwięziony i powoduje "globalne ocieplenie". Naukowcy spodziewają się, że w ciągu najbliższych 100 lat temperatury mogą wzrosnąć do 10 stopni Fahrenheita, a poziom morza może wzrosnąć nawet o 20 cali. Za zaledwie sto lat świat będzie wyglądał zupełnie inaczej niż dzisiaj. Polarne czapy lodowe topnieją i zagrażają bytowi dużych ssaków, takich jak niedźwiedzie polarne, których strategia polowania opiera się na pływającym lodzie w celu łapania ofiar, takich jak foki, w wodzie pod nimi. Bardzo niewielkie zmiany temperatury wpływają na mniejsze zwierzęta, które nie są w stanie przystosować się ani poruszać w klimacie. Na przykład gatunki rzekotek drzewnych wymierają, ponieważ te, które żyją na szczytach górskich, stale zbliżają się do szczytu, aż nie mogą już znaleźć niższych temperatur. Ludzie mają historię powodowania wymierania zwierząt lub przynajmniej odpowiedzialności za nie. Na przykład za znikanie leniwców naziemnych w Indiach Zachodnich i mamutów karłowatych na Wyspie Wrangla na Oceanie Arktycznym obwinia się ludzkich myśliwych. Wymieranie małych i szczególnie dużych zwierząt (megafauna) nastąpiło w obu Amerykach między 12 a 10 Kya, na Madagaskarze między 6 a 1 Kya, a w Australii między 40 a 30 Kya i nadal jest dyskutowane, czy zostały wywołane przybyciem ludzi do regionów lub jeśli były stymulowane przez zmiany klimatyczne. W Ameryce Północnej megafauna (w tym mamuty, mastodonty, gigantyczne bobry, dzikie konie, wielbłądy i koty szablozębne) zniknęła z zapisów kopalnych mniej więcej w tym samym czasie, gdy pojawiły się niektóre z najwcześniejszych artefaktów, takich jak lud Clovis. Krytycy twierdzą, że najlepszym dowodem na to, że nadmierne polowanie na ludzi spowodowało wymieranie, byłoby obfitość miejsc zabicia (poćwiartowane szkielety z grotami włóczni), ale nie ma ich zbyt wiele. Możliwe jednak, że zmiana klimatu spowodowana regularnymi cyklami glacjalnymi w połączeniu z groźbą sprytnego ludzkiego drapieżnika mogła być śmiertelnym duetem. Być może poprzez nadmierne zabijanie pasących się zwierząt ludzie pośrednio spowodowali zmiany środowiskowe w krajobrazie, co wpłynęło na inne zwierzęta. Obecnie przyjazne dla środowiska wzorce polowań są zarządzane przez lokalne, stanowe i federalne instytucje rządowe w Stanach Zjednoczonych. Jednak przepisy dotyczące polowań i połowów mogą nie wystarczyć, aby powstrzymać nas przed powtórzeniem naszych błędów z przeszłości. Z powodu przełowienia i zanieczyszczenia naukowcy przewidzieli, że jeśli nie zostaną wprowadzone drastyczne zmiany, w ciągu najbliższych pięćdziesięciu lat prawie wszystkie morskie gatunki owoców morza ulegną załamaniu. Jak na ironię, ludzie mogą być odpowiedzialni za zabicie jednego z kluczowych źródeł żywności, które przyczyniły się do uczynienia nas "ludźmi", ponieważ włączenie zwierząt morskich do naszej paleolitycznej diety mogło wpłynąć na ewolucję naszego mózgu.
Początki Człowieków (65)
EWOLUUJEMY CZY WYGAŚNIEMY?
Istnieją opowieści zrodzone z niezrozumienia ewolucji, że ludzie w końcu stracą swój maleńki piąty palec u nogi lub pękną wyrostki. Przewiduje się, że mężczyźni stracą swoje pozbawione mleka sutki, a kobiety rozwiną bardzo duże lub nawet dodatkowe piersi, aby dostosować się do przesadnych preferencji seksualnych. Powszechnie zakłada się nawet, że populacja na świecie w końcu będzie miała jeden odcień skóry, mimo że zarówno z doświadczenia życiowego, jak i z genetyki wiemy, że dziedziczenie koloru skóry nie jest tak proste, jak mieszanie ze sobą pigmentów każdego z rodziców. Te spekulacje w naturalny sposób prowadzą do pytania: czy dobór naturalny już na nas działa? Z przykładów takich jak tolerancja na laktozę wiemy, że ewolucja spowodowała poważne zmiany w populacjach ludzkich w ciągu zaledwie kilku tysięcy lat. W niektórych populacjach odporność na zakażenie wirusem HIV i malarią jest wyższa niż w innych. Wyraźnie ludzkie adaptacje kulturowe i innowacje nie powstrzymują nas przed rozwojem niektórych biologicznych. Ale jeśli dobór naturalny działa obecnie w populacji ludzkich, czy to oznacza, że na horyzoncie pojawia się specjacja? I czy możemy odważyć się zapytać o coś takiego bez flirtowania z rasizmem? Przy nowoczesnych sposobach podróżowania po świecie i zmniejszających się barierach kulturowych między populacjami (języki wymierają na całym świecie), przepływ genów najprawdopodobniej zapobiegnie rozgałęzianiu się nowych gatunków od ludzi. Przy 7 miliardach nas na planecie wydaje się mało prawdopodobne, aby jakakolwiek populacja ludzka mogła zostać tak reprodukcyjnie izolowana, że dzieli się na oddzielny gatunek, ale taki scenariusz nie wykracza poza granice rzeczywistości, zwłaszcza w następstwie hipotetycznej ogólnoświatowej katastrofy jak doświadczane przez dinozaury. Na szczęście NASA opracowuje plany odwrócenia asteroidy, takiej jak ta, która spowodowała wyginięcie dinozaurów w przypadku ponownego zagrożenia Ziemi. Pomijając katastrofy, których można uniknąć lub których nie da się uniknąć, czy możemy użyć leków, aby nie wyginąć? Medycyna wraz z technologią rolniczą prawdopodobnie pomogą powstrzymać H. sapiens przed przewidywalnym zgonem, ale nie mogą pomóc nam ewoluować w formy bardziej odporne na wymieranie. Dopóki nie zostaną wprowadzone medyczne zmiany w gametach i nie zostaną odziedziczone w przyszłych pokoleniach, wszystkie najlepsze terapie odrastania narządów i przeciwdziałania starzeniu się będą prawie nic nie znaczyły w kategoriach ewolucyjnych. Ale dobór naturalny może nadal działać na płodność lub przeżycie płodu, a dobór płciowy również może zmienić sytuację. Istnieją dowody na to, że wiele naszych genów zaangażowanych w tolerancję laktozy, rozwój mózgu, pigmentację skóry, rozwój narządów rozrodczych, metabolizm i odporność na choroby przechodzi obecnie silną selekcję, która najprawdopodobniej doprowadzi do zmian przynajmniej na poziomie populacji w przyszły. Jako gatunek będziemy nadal ewoluować w przyszłość, ale jak bardzo? Moglibyśmy ewoluować do form tak odmiennych od naszych obecnych, że przyszli biolodzy i paleontolodzy nazwaliby nas innym gatunkiem. W tym sensie H. sapiens, jaki znamy teraz, wyginąłby. Możemy jednak nie mieć dużo więcej czasu na ewolucję, ponieważ przewiduje się, że następny duży obrót fauny lub cykliczne wymieranie nastąpi około 500 000 n.e. Przeciętnie gatunki ssaków trwają około 2,5 miliona lat, co jest okresem między zdarzeniami obrotu. To prawdopodobnie nie przypadek, że drastyczna ewolucja linii homininów odpowiada ostatniemu zdarzeniu obrotu około 2 milionów lat temu. Te wymierania są rozdzielone interwałami zmian klimatu (wpływających na temperatury, opady, siedliska i źródła pożywienia), które są kontrolowane przez astronomiczne zmiany, takie jak zmiany orbity Ziemi i jej chybotanie się na jej osi (cykle Milankovitcha). 500 000 lat to mnóstwo czasu na ewolucję w coś nowego, w końcu 500 Kya na Ziemi byli tylko średnio inteligentni Archaicy i H. erectus i spójrzcie, gdzie dziś stoją homininy. Ale podczas gdy nasza populacja liczy 7 miliardów, nasi najbliżsi krewni są na skraju wyginięcia. World Wildlife Fund (www.worldwildlife.org) donosi, że wszystkie wielkie małpy człekokształtne są zagrożone. Na wolności pozostało tylko około 10 000 bonobo, w porównaniu z ponad 100 000 zaledwie dwadzieścia lat temu. Są one wyciskane z preferowanych siedlisk przez ludzi, a także zabijane, aby mięso z buszu było podawane w drogich restauracjach. Orangutany utknęły na wyspach, które są coraz bardziej wylesiane. Ich liczba spada do 50 000. Jane Goodall poinformowała na swojej stronie internetowej (www.janegoodall.org), że w 2004 roku pozostało tylko 150 000 szympansów, ale dziś powinno ich być co najmniej milion. Do dnia dzisiejszego przetrwało mniej niż 1000 goryli górskich bez pomocy kłusowników i śmiertelnych epidemii wirusa Ebola. Świadek nieuniknionego dzikiego wyginięcia naszych najbliższych krewnych przypomina nam, jak bardzo potrafimy się przystosować, ale to, że zwykle opowiadamy jedną historię ewolucyjną (naszą własną), nie oznacza, że cała ewolucja kulminowała w wyniku działalności człowieka. Chociaż dla wielu ludzi bycie człowiekiem oznacza wyniesienie się z ryb i reszty królestwa zwierząt, samo decydowanie, że ludzie są królem planety, nie oznacza, że tak naprawdę nimi jesteśmy. To prawda, że ludzie pokrywają prawie całą suchą powierzchnię świata, a także część mokrej powierzchni, ale po zaledwie 6 milionach lat ewolucji żyje tylko jeden gatunek małp człekokształtnych, a mimo to istnieje ponad 350 000 gatunków (i wciąż rośnie). chrząszczy, które ewoluują od ponad 260 milionów lat. Pod tym względem chrząszcze są królem planety, z powodzeniem promieniując i rozmnażając się znacznie częściej niż ludzie kiedykolwiek mieli i kiedykolwiek będą.
Początki Człowieków (64)
Czy neandertalczycy odkryli Amerykę?
Podczas przesiewania brudu i skał z koryta potoku w poszukiwaniu śladów Kanału Santee zbudowanego w latach 90. XVIII wieku (30 mil na zachód od Charleston w Południowej Karolinie), podwodny archeolog Mark Newell znalazł surowe narzędzie z kamienia. Było to niezwykłe znalezisko, biorąc pod uwagę, że narzędzie to zostało wykonane przez neandertalczyków w Europie. Dodatkowo surowiec był oczywiście obcy. Mikroskopijne skamieniałości bezkręgowców w skale potwierdziły, że pochodzi ona z Francji. W rzeczywistości unikalny skład geochemiczny kamienia wskazywał na jego źródło w konkretnej formacji skalnej w regionie Bordeaux. Czy neandertalczycy mogli przepłynąć Atlantyk w epoce kamienia, aby odkryć Amerykę Północną? Kuszące byłoby stwierdzenie, że neandertalczycy byli pierwszymi Amerykanami, ale po dalszej ocenie proste wyjaśnienie staje się jasne. W stosunkowo niedawnej historii europejskie rzeki, w których znajdują się prehistoryczne artefakty, były wydobywane w celu uzyskania balastu, a następnie europejskie statki zrzucały swój balast w amerykańskich portach, zabierając ładunek na podróż powrotną do domu. Te wysypiska były następnie wydobywane w celu uzyskania balastu, który został poddany recyklingowi w lokalnych amerykańskich jednostkach pływających, takich jak te, które pływały w kanale Santee.
Początki Człowieków (63)
ROZPROSZENIE NA CAŁYM ŚWIECIE
Zaludnienie planety należy traktować raczej jako konsekwencję rozproszenia niż zamierzonej migracji. Należy ją sobie wyobrazić jako płynny proces, w którym ludzie poruszają się tam i z powrotem, w przeciwieństwie do pojedynczego, ukierunkowanego zdarzenia kolonizacji. Przyczyna początkowego rozproszenia homininów z Afryki nie jest wyraźnie określona w skamielinach ani w zapisach archeologicznych. Wczesna hipoteza sugerowała, że wynalezienie siekiery aszelskiej umożliwiło zasiedlenie H. erectus nowego terytorium. W pewnym sensie może pozwolić sobie na odważne wyjście z nową technologią. Pomysł opiera się na powiązaniu wszechobecnego siekiery ręcznej z H. erectus i obecności siekier ręcznych poza Afryką. Założenia stojące za tym pomysłem są zbyt jasne dla większości dzisiejszych naukowców, biorąc pod uwagę słonie rozproszone z Afryki bez korzystania z siekier ręcznych. Coś bardziej biologicznego niż kulturowego było prawdopodobnie przyczyną rozprzestrzeniania się H. erectus. Gatunek drapieżny nie może przetrwać w dużych grupach. Ponieważ znajduje się na szczycie piramidy troficznej, drapieżnik nie może przewyższyć swojej ofiary lub umrze z głodu. Pat Shipman i Alan Walker zasugerowali, że aby H. erectus mógł dokonać przemiany w mięsożercę, prawdopodobnie albo zmniejszył liczebność populacji, albo zwiększył zasięg geograficzny, a następnie zwiększył rozmiar ciała. Zapis kopalny wskazuje, że te dwa ostatnie wyraźnie pojawiły się po 1,8 miliona lat temu, mniej więcej wtedy, gdy dowody na jedzenie mięsa stają się powszechne. Rozproszenie H. erectus w Starym Świecie było prawdopodobnie konsekwencją stania się drapieżnikiem i po prostu podążania za stadami. Przyczyna rozproszenia się współczesnych ludzi mogła być wynikiem podobnych nacisków. Drapieżniki były z pewnością problemem dla wczesnych ludzi, jednak wraz z pojawieniem się H. sapiens mamy pierwsze wyraźne dowody na ogień, schronienie, wyższy stopień uspołecznienia, wzrost wielkości mózgu, poprawę umiejętności technologicznych. Ewolucja technologii i zdolności adaptacyjnych prawdopodobnie ułatwiły ekspansję ludzi do bardziej marginalnych i wcześniej niedostępnych środowisk w porównaniu z H. erectus. Istnieje również wiele dowodów na to, że współcześni ludzie w różnych regionach włączali do swojej diety różne ofiary, takie jak żółwie, zające, ptaki naziemne i skorupiaki, więc niekoniecznie wszystkie populacje podążały za stadami. Miejsca wzdłuż wybrzeży świata sugerują, że wczesne populacje ludzkie z powodzeniem żyły na wybrzeżach, a ten styl życia prawdopodobnie ułatwiał rozproszenie, zwłaszcza w czasach, gdy łodzie były wszechobecne. Pochodzenie łodzi i statków morskich jest obecnie niejasne, ale ogólnie można założyć, że pierwsi Australijczycy w wieku od 60 do 50 Kya mieli takie wynalazki. Kolonizacja Australii jest intrygująca, ponieważ nawet przy niskim poziomie morza nigdy nie było połączenia lądowego między szelfem kontynentalnym Sunda w północno-zachodniej Indonezji a szelfem kontynentalnym Sahul, który zawiera ląd australijski, a także Papuę-Nową Gwineę i Tasmanię. Australia jest niezwykłym przypadkiem, ponieważ kontynent mógł nie być widoczny z żadnej indonezyjskiej wyspy, nawet w czasach, gdy poziom morza był najniższy w okresach lodowcowych, kiedy woda jest związana na zamarzniętych biegunach. Chociaż schematy lotu ptaków mogły wskazywać ląd poza horyzontem, odległość jest uważana za zbyt dużą, aby ludzie mogli po prostu przeskoczyć na wyspę, tak jak zrobił to H. erectus (i być może Homo floresiensis) w całej Indonezji. Zakłada się, że aby dostać się do Australii, ludzie musieliby zbudować łodzie lub prymitywne tratwy i wyruszyć na otwarte morze w celu eksploracji. Dotarcie do Australii prawdopodobnie wymagało technicznych umiejętności żeglarskich, a także umiejętności planowania, pracy w grupie i prawdopodobnie rozmawiania ze sobą. Po dotarciu na dużą wyspę istnieją dowody na to, że szybko przenieśli się na południową część. Do 23 Kya w Willandra Lakes w południowo-wschodniej Australii pierwsi mieszkańcy pozostawili po sobie największą na świecie kolekcję ludzkich śladów plejstocenu. Skamieniałe czaszki i szkielety z najwcześniejszych miejsc, takich jak Bagno Kow w północnej Wiktorii i jezioro Mungo, pokazują, że tylko współcześni ludzie byli w stanie dotrzeć do Australii. Pierwsi Amerykanie byli też prawdopodobnie ludźmi żeglarzami. Pochodzili z północno-wschodniej Azji i byli nie tylko dobrzy w morskim stylu życia, ale także przystosowali się do arktycznego. Analizy genetyczne szacują, że ludzie po raz pierwszy przybyli do obu Ameryk już w wieku 30 Kya, ale najprawdopodobniej między 25 a 15 Kya w szczytowym okresie ostatniego zlodowacenia. Najbardziej prawdopodobna trasa przebiegała przez Cieśninę Beringa z Syberii na Alaskę, która przy niższych poziomach mórz byłaby w dużej mierze zamknięta, a niektóre wyspy aleuckie zostałyby połączone, dzięki czemu podróż (choć niekoniecznie zamierzona) byłaby bardziej zakotwiczona na lądzie i dlatego mniej zdradzieckie. Jon Erlandson sugeruje, że pierwsi Amerykanie mogli wydostać się z Azji do Ameryki "starożytną autostradą z wodorostów". Zgodnie z "teorią migracji przybrzeżnej" mogli żyć w małych populacjach morskich, które pływały z wyspy na wyspę lub z brzegu na brzeg, polując na stworzenia morskie żyjące w lasach wodorostów. Obecnie istnieje prawie ciągła autostrada z wodorostów z Japonii i Syberii, przez Cieśninę Beringa na Alaskę i wzdłuż wybrzeża Kalifornii. A w lasach wodorostów żyją na przykład foki, wydry morskie, ryby i jeżowce. Niektóre z najwcześniejszych stanowisk archeologicznych w obu Amerykach znajdują się w pobliżu produktywnych lasów wodorostów. Daisy Cave na Wyspach Normandzkich w południowej Kalifornii, datowana na prawie 10 Kya, zachowała dowody na to, że ludzie używali tam zasobów wodorostów. Kultura Clovis, nazwana tak od odkrycia w Clovis w stanie Nowy Meksyk, znana jest z elegancko wyróżniających się grotów włóczni i datuje się na 11 Kya pod koniec ostatniej epoki lodowcowej. Ludowi Clovis od dawna nadano tytuł najwcześniejszych ludzi, którzy przybyli do Ameryki, ale geografia większości stanowisk jest przekrzywiona na wschodnią część Ameryki Północnej, podczas gdy wcześniejsze i współczesne miejsca zajmowane przez ludzi przed Clovisem znaleziono na zachodzie oraz w Ameryce Południowej. Duktai na Alasce ma około 12 Kya, a inne, takie jak schronisko skalne Meadowcroft w Pensylwanii, Topper w Południowej Karolinie, Taima-Taima w Wenezueli, Pedra Furada w Brazylii i Monte Verde w Chile są równie stare i mogą być nawet starsze. Zwłaszcza Meadowcroft może mieć nawet 20 Kya. Około godziny 12.5 Kya w Monte Verde w Chile archeolodzy znaleźli najwcześniejszą stałą osadę w Nowym Świecie z pozostałościami schronu, który byłby wystarczająco duży, aby pomieścić ponad trzydzieści osób. Podobnie jak w przypadku rozproszenia się z Afryki, prawdopodobnie nastąpił wielokrotny napływ ludzkich populacji do Nowego Świata, a ciągłe odkrywanie stanowisk archeologicznych, a także głębsza analiza genetyki żywych populacji ludzkich pomoże opowiedzieć prawdziwą historię pierwszych Amerykanów.
Początki Człowieków (62)
Wyścig
H. sapiens to jeden zmienny gatunek, który utrzymuje się razem dzięki przepływowi genów na dużą skalę. Pomiędzy populacjami o kontrastujących częstościach alleli istnieją populacje o pośrednich częstościach. Zjawisko to jest znane jako klin lub gradient częstotliwości alleli w przestrzeni utrzymywany razem przez przepływ genów. Zmienność ludzka reprezentowana przez linię jest ciągła w widmie i nie ma odrębnych grup. Dlatego pojęcie "rasy" używane dzisiaj zostało uznane za strasznie błędne i powinno zostać zastąpione pojęciem ciągłej różnorodności geograficznej w biologii i kulturze. Chociaż współczesna naukowa mantra mówiąca, że "nie ma biologicznych podstaw dla rasy" jest prawdziwa na wielu poziomach, jest również myląca. Oczywiście istnieją regionalne różnice fenotypowe między populacjami ludzi i są one widoczne na przykład w kolorze skóry, kolorze i strukturze włosów, kolorze i kształcie oczu, kształcie nosa, wzroście, długości kończyn i proporcjach ciała, kształcie czaszki i twarzy, uchu typ wosku, grupa krwi, podatność i odporność na choroby, tkanka tłuszczowa dystrybucja, adaptacje na dużych wysokościach, tolerancja laktozy i tak dalej. Różnice te są spowodowane przypadkowym dryfem genetycznym, selekcją płciową, różnymi poziomami przepływu genów i oczywiście regionalnymi adaptacjami środowiskowymi. Zmienność regionalna rejestruje również historię przodków. Kształt oczu i tekstura włosów rdzennych Indian amerykańskich odzwierciedla ich azjatyckie pochodzenie. Cechy, których używamy do identyfikacji "ras", są prawie zawsze cechami ciągłymi, które występują w różnym stopniu we wszystkich populacjach ludzkich. Badania nuklearnego DNA dotyczące zmienności geograficznej wykazały, że cechy, które odzwierciedlają zmienność regionalną, są determinowane przez niewielki ułamek naszych genów. Co więcej, niespełna 15 procent ludzkiej zmienności genetycznej oddziela się główne grupy ludzkie. W porównaniu z różnymi gatunkami dużych ssaków o doskonałych zdolnościach rozpraszania (np. wilk szary i jeleń wirginijski), ludzie mają bardzo małą zmienność. Większość różnorodności ludzkiej istnieje jako różnice między jednostkami w obrębie populacji. Oznacza to, że w populacjach występuje większe zróżnicowanie niż między nimi.
Początki Człowieków (61)
MODELE WIELOREGIONALNE I POZA AFRYKĄ
Homo erectus był pierwszym homininem, który rozprzestrzenił się poza Afrykę. Do tego czasu wszystkie skamieniałości homininów od około 6 do 2 milionów lat temu są ograniczone do Afryki. Prawie jak tylko H. erectus pojawił się na sawannach Afryki Wschodniej około 1,8 mln lat temu, rozprzestrzenił się na północ aż do Dmanisi w Gruzji i na wschód do Jawy w Indonezji. H. erectus przetrwał w niektórych częściach Indonezji aż do 30 Kya. Ten ogromny zasięg geograficzny i czasowy H. erectus jest inspiracją dla dwóch głównych rywalizujących teorii współczesnego pochodzenia geograficznego i rozproszenia człowieka. Chociaż hipotezy wielu paleoantropologów znajdują się gdzieś pośrodku, te dwa modele początków współczesnego człowieka są znane jako model "poza Afryką" (OD) lub "zastąpienie" lub "ogród Eden" oraz model wieloregionalny (MR) lub "ciągłość regionalna". lub model "Krata". Pomimo nazw modeli, obaj zgadzają się, że korzenie ewolucji człowieka leżą w Afryce. Po prostu nie zgadzają się co do czasu i wielkości udziału ewolucyjnego populacji H. erectus i archaicznych na całym świecie. Obecnie coraz więcej naukowców opowiada się za OA ze względu na przekonujące dowody genetyczne, ale żaden model nie został ostatecznie poparty z wykluczeniem rywala, ani nie został ostatecznie obalony. Ścisły model MR, wspierany przez Milforda Wolpoffa i innych, opiera się na hipotezie, że homininy, które najpierw rozproszyły się z Afryki, wyewoluowały w różnych regionach geograficznych, a następnie krzyżowały się ze współczesnymi ludźmi, rozprzestrzeniając się po Starym Świecie. Innymi słowy, H. erectus i archaiczni ludzie (np. neandertalczycy i Homo heidelbergensis) rozprzestrzenili się do Europy, Azji, Indonezji, a następnie w odpowiednich regionach wyewoluowali w Homo sapiens. W MR populacje w różnych regionach geograficznych były powiązane z przepływem genów, tak jak jest dzisiaj, ale przepływ genów zaczął się 1,8 miliona lat temu. Dowody potwierdzające tę hipotezę są głównie morfologiczne. MR wskazuje, że podobnie jak współcześni ludzie, wymarłe homininy wykazują zmienność geograficzną i adaptacje, a zwolennicy łączą tę regionalną anatomię homininów ze współczesnymi ludźmi żyjącymi obecnie w tych regionach. Współczesne populacje chińskie zachowują cechy podobne do skamieniałego H. erectus i archaicznych ludzi z tego regionu, takie jak siekacze w kształcie łopaty oraz pewne cechy kształtu twarzy i czaszki. Dokonuje się również porównań między H. erectus a rdzennymi Australijczykami. Uważa się również, że współcześni Europejczycy zachowują podobieństwa do neandertalczyków, takie jak rozmiar nosa, rozmiar ciała, postura i solidność. Kuszące dowody na krzyżowanie się neandertalczyków ze współczesnymi ludźmi również wspierają MR. Odkrywcy szkieletu dziecka w Lagar Velho w Portugalii sugerują, że jest to "dziecko miłości" ze względu na mieszankę cech neandertalczyków i współczesnych ludzi. Szkielet pochowany z przebitymi muszlami i czerwoną ochrą z Lagar Velho w Portugalii pochodzi z około 25 Kya i ma neandertalskie proporcje kończyn z ludzkim podbródkiem. Krytycy nie są jednak przekonani do twierdzenia o hybrydach i twierdzą, że proporcje mieszczą się w zakresie ludzkim, choć na końcu spektrum przystosowanego do zimna. Erik Trinkaus widzi cechy mozaiki również w innych europejskich skamielinach, na przykład z Rumunii około 40 Kya. Niestety, nie ma sposobu na udowodnienie, że neandertalczycy i współcześni ludzie nie krzyżowali się bez znalezienia np. szkieletu neandertalczyka pochowanego w tym samym grobie, co współczesny człowiek. Być może w miarę powiększania się mapy genomu neandertalczyka i rozumienia funkcji genów wzrostu, genetycy będą w stanie określić, czy istniałyby bariery molekularne w ich krzyżowaniu się z ludźmi. Ścisły model OA utrzymuje, że po tym, jak homininy rozproszyły się najpierw z Afryki jako H. erectus i Archaics, drugie główne rozproszenie współczesnych ludzi zastąpiło istniejące populacje na całym świecie bez mieszania genetycznego. (Chociaż luźniejsze interpretacje OA pozwalają na niewielki przepływ genów). Zastąpienie mogło być wynikiem pośredniej konkurencji, takiej jak po prostu przetrwanie bardziej prymitywnych grup, lub bezpośredniej konkurencji, takiej jak wojny i ludobójstwo. Ścisła OA nie obejmuje krzyżowania się współczesnych ludzi i archaików i twierdzi, że różnorodność geograficzna współczesnego człowieka pojawiła się niedawno po 200 Kya. Skamieniałości i dowody archeologiczne potwierdzają OA, ponieważ najstarsi współcześni ludzie występują w Afryce (Omo I i II). Podobieństwa anatomiczne we współczesnych populacjach ludzkich są niezmiernie bliższe niż podobieństwa między ludźmi przednowoczesnymi i współczesnymi w danym regionie geograficznym. Co więcej, gdyby archaicy ewoluowali w nowoczesnych ludzi, ich istnienie nie powinno się pokrywać, jednak w izraelskich miejscach Skhul i Qafzeh od 110 do 90 Kya współcześni ludzie żyli bardzo blisko neandertalczyków w pobliskim miejscu Tabun około 110 Kya. Pierwsi współcześni ludzie przybyli do Europy przed 40 Kya i mieli tropikalne adaptacje w swoich proporcjach ciała, ale neandertalczycy nadal ewoluowali tam do 28 Kya z ich adaptacjami do zimna. Po 40 Kya istnieje wiele nowoczesnych miejsc pracy dla ludzi w całej Europie Zachodniej. Najstarsze dowody pochodzą ze stanowisk takich jak Bacho Kiro i Temnata w Bułgarii (43-40 Kya), Pestera cuoase w Rumunii lub "jaskinia z kośćmi" (40-35 Kya), a następnie w Kent&prim;s Cavern w Anglii (30 Kya) . Nakładanie się współczesnych ludzi i neandertalczyków w Europie trwało około 10 000 lat przed zniknięciem neandertalczyków. Niektórzy zwolennicy OA obwiniali ludzi o spowodowanie śmierci neandertalczyków. W miarę gromadzenia odkryć na Bliskim Wschodzie i w Europie wyłonił się wyraźny wzór: stanowiska współczesnego człowieka i neandertalczyków pozostały odrębne, ale stanowiska neandertalskie z biegiem czasu stawały się coraz mniej i bardziej od siebie oddalone. Większość dotychczasowych dowodów morfologicznych dotyczących modelu MR nie jest tak przekonująca, jak dowody molekularne, które wspierają model OA. Rozproszenie współczesnych ludzi z Afryki Subsaharyjskiej nastąpiło po 200 Kya, zarówno według analiz mitochondrialnego DNA, jak i chromosomu Y. Dowody starożytnego DNA (aDNA) również wspierają OA. DNA neandertalczyka leży poza normalnym zakresem zmienności występującym u współczesnych ludzi, ale nie jest tak rozbieżne, jak sekwencje szympansów u współczesnych ludzi. Dystans genetyczny między neandertalczykami a współczesnymi ludźmi, taki sam nawet dla współczesnych Europejczyków, sugeruje, że neandertalczycy nie wyewoluowali w ludzi nowoczesnych ani nie przyczynili się w sposób od przodków do potomków do początków współczesnego człowieka. Nawet aDNA ze skamieniałości, które mają to, co niektórzy badacze nazywają anatomią przejściową, nie jest przejściowe.
Początki Człowieków (60)
WOJNA CZY POKÓJ?
W przeciwieństwie do innych naczelnych, możliwe jest, że ludzie żyją w grupach ze względu na potrzebę ochrony drapieżników przed przedstawicielami własnego gatunku. Chociaż inne naczelne okazują przemoc wobec własnego gatunku, ludzie są wyjątkowo dobrzy w zabijaniu się nawzajem. Z pewnością, gdy doprowadzeni do skrajności, jak w czasie suszy lub na wyspach, gdzie zasoby są szybko przewyższane, ludzie zaatakują i zjedzą własne gatunki. Liczne stanowiska archeologiczne na całym świecie mają mocne dowody na ludzki kanibalizm. Nawet stanowisko neandertalczyków 43 Kya w Hiszpanii w jaskini El Sidr′on pokazuje, że kilka (do ośmiu) szkieletów, które były już ofiarami złego stanu zdrowia i złego odżywiania (co widać po liniach stresu zwanych "hipoplazjami" w szkliwie zębów ), zarżnięto, rozczłonkowano i zjedzono. Kanibalizm nie jest rzadkością pod przymusem i jest powszechnym zjawiskiem wśród zwierząt. Wojny i ludobójstwa są jednak rzadkością, ale czy możemy winić za to naszych przodków? Dzielimy historię ewolucji z naczelnymi, które praktykują dzieciobójstwo jako udaną adaptację. Samce goryli i langurów szarych (małpy Starego Świata), które są nowe w grupie samic, zabijają niemowlęta, aby jak najszybciej mogły rozpocząć własną linię i zwiększyć swój własny sukces reprodukcyjny. Nasi jeszcze bliżsi krewni, szympansy zwyczajne, są wysoce terytorialne. Samce patrolują obrzeża i granice i atakują, a czasem zabijają członków sąsiednich grup. Społeczeństwo szympansów charakteryzuje się hierarchicznymi związkami mężczyzn, które różnią się od bonobo pomimo ich niedawnego podziału ewolucyjnego nieco ponad 2 miliony lat temu. Podczas gdy szympansy rozwiązują konflikty za pomocą polityki i przemocy, bonobo (szympansy karłowate) pogodzą się z miłością i seksem. Mają społeczeństwo bardziej zorientowane na kobiety, w którym samice współpracują w przeciwieństwie do szympansów. Bonobo witają rywalizujące grupy uściskami dłoni na genitaliach i zmysłowymi maseczkami ciała, aby uniknąć konfliktu, a jeśli wystąpi konflikt, zostanie on szybko rozwiązany za pomocą pocałunków i seksu w różnych pozycjach między mężczyznami i kobietami, kobietami i kobietami oraz mężczyznami i mężczyznami. Czy istnieje sposób, aby dowiedzieć się, który gatunek najlepiej przybliża LCA? Takie ćwiczenia umysłowe należy wykonywać ostrożnie, ponieważ o małych populacjach bonobo żyjących na wolności wiadomo znacznie mniej niż o szympansach. Wraz z rosnącymi obserwacjami bonobo w terenie staje się coraz bardziej jasne, że oba gatunki są bardziej podobne, niż nam się kiedyś wydawało. Do tej pory większość pokojowych zachowań bonobo znano tylko z niewoli, gdzie zawsze było pod dostatkiem pożywienia. Co więcej, nasza tendencja do identyfikowania się z bonobo z powodu ich rzekomej tendencji do zachowania dwunożnego jest teraz kwestionowana, ponieważ niektórzy naukowcy, tacy jak Bill McGrew, wykazali, że bonobo i szympansy wykonują równoważne ataki dwunożności. Dowody wojny lub pokoju z dawnych czasów w prehistorycznych zapisach były interpretowane na różne sposoby. Pracując codziennie wśród skamieniałości, Raymond Dart zrozumiał, ile części szkieletu byłoby dobrą bronią. Szczęki krzywokształtne z postrzępionymi zębami do cięcia, duże, ciężkie kości przeznaczone do tępych sił, złamane kości długie, które łuszczą się jak kamień i mogą być ostre jak kamień, to wszystko części, które mogą i mogłyby wyrządzić poważną krzywdę. Obfitość pozornie wspaniałej broni w jaskini Swartkrans w Południowej Afryce doprowadziła Darta, we wczesnych latach, w pierwszej połowie XX wieku, do spekulacji, że homininy, które tam żyły, miały agresywną kulturę. Jeśli nie po to, by zabić ofiarę, używali jej kości do grożenia, okaleczania lub zabijania się nawzajem. Hipoteza ta stała się znana jako hipoteza "morderczej małpy", a ich zestaw narzędzi nazwano kulturą "osteodontokeratyczną". Pomysł stracił poparcie przez lata wraz z rosnącym zrozumieniem tafonomii. Zabierz błędnie zinterpretowaną broń, a australopity nie były szczególnie groźne. Choć mogły być równie silne jak szympansy, miały znacznie mniejsze zęby. Co więcej, nie ma bezpośrednich dowodów na to, że robili kamienne narzędzia do 2,5 miliona lat temu (jeśli były to gatunki, które je wytwarzały). Pozbawione ostrych zębów, rozmiarów i zaawansowanej technologicznie broni australopity prawdopodobnie wykorzystywały mózg, zwinność i umiejętności społeczne, aby uciec przed drapieżnikami. Kiedy wyewoluowało rzucanie przez ramię i możliwe było działanie na odległość, homininy mogły nie tylko odpędzać drapieżniki i konkurentów za mięso, ale oszuści (we wzajemnych stosunkach) mogą zostać ukarani niewielkim kosztem dla ukaranego (w przeciwieństwie do walki wręcz). Rzucanie bez wątpienia pozwalało na prowadzenie działań wojennych, a kiedyś technologia narzędziowa, taka jak miotacze włóczni wynalezionych przez 30 Kya, zwiększała odległość i siłę wywieraną na pociski.
Początki Człowieków (59)
ALTRUIZM I KOLONIA LUDZKA
Życzliwość niekoniecznie jest cechą wyłącznie ludzką, ale ludzie wydają się szczególnie skłonni do bezinteresownych czynów na własny koszt. Dobór naturalny przewiduje ewolucję cech kooperatywnych (korzyść dla wszystkich) i egoistycznych (korzyść dla jednostki, koszt dla kogoś innego), ponieważ obie zwiększają przystosowanie jednostki, która je nosi. Altruistyczne (koszt dla jednostki, korzyść dla kogoś innego) i złośliwe (koszt dla wszystkich) obniżają przystosowanie ich nosicieli i przewiduje się, że dobór naturalny nie będzie sprzyjał tym zachowaniom. Tak więc altruizm jest ewolucyjną zagadką; jest to zachowanie, które wydaje się niezgodne z doborem naturalnym. W jaki sposób dobór może sprzyjać kosztownym zachowaniom, zwłaszcza takim, które utrudniają lub faktycznie uniemożliwiają jednostce pomyślne rozmnażanie się? Zjawisko altruizmu jest udokumentowane u wielu zwierząt, zwłaszcza u owadów, takich jak pszczoły robotnice. Psy myśliwskie i sójki z Florydy to tylko dwa gatunki, w których znaczna liczba dorosłych nie rozmnaża się, a zamiast tego pomagają innym wychowywać więcej potomstwa, niż mogliby sobie poradzić sami. Teoria doboru grupowego przewiduje, że cechy altruistyczne powinny się rozprzestrzeniać. Geny altruizmu będą utrwalane w przyszłych pokoleniach, ponieważ altruizm jest dobry dla grupy jako całości. (Geny altruizmu niekoniecznie bezpośrednio powodują, że jednostka zachowuje się bezinteresownie, mogą na przykład kodować satysfakcjonujący, poprawiający samopoczucie naturalny haj dopaminy lub środek redukujący stres, który włącza się, gdy dana osoba zachowuje się altruistycznie). ta logika polega na tym, że cecha będzie się rozprzestrzeniać tylko wtedy, gdy geny, które ją kształtują, są dobre w dostaniu się do następnego pokolenia. Geny altruizmu są złe w dostaniu się do przyszłych pokoleń. Co więcej, jednostki altruistyczne zwiększają sprawność konkurujących jednostek, ponieważ zmniejszają swoją własną. Zgodnie z teorią doboru grupowego altruiści byliby wybierani przeciwko. Ogólnie rzecz biorąc, korzyści dla grupy na ogół nie wystarczają do rozpowszechnienia cechy. Połączenie doboru naturalnego i doboru krewniaczego rozwiązuje zagadkę altruizmu, przynajmniej gdy krewni odnoszą korzyści. Pokrewne dzielą geny, więc selekcja powinna faworyzować altruizm, który jest wybiórczo nakierowany na krewnych. W tym scenariuszu, przedstawionym przez W.D. Hamiltona w 1964 roku, egoizm byłby nieprzychylny, a współpraca byłaby faworyzowana. Altruista może mieć niskie dopasowanie bezpośrednie, ale jego ogólna lub "włączająca" sprawność byłaby wysoka, gdyby pomogli w udanym przetrwaniu i reprodukcji swoich krewnych, którzy dzielą geny. Wzajemność jest jednak kluczem do altruizmu niespokrewnionego. Mówiąc dokładniej, wzajemny altruizm, koncepcja wprowadzona przez Roberta Triversa w 1971 roku, może wyjaśniać zachowanie, gdy spełnione są dwa warunki: 1) koszty muszą być niewielkie w porównaniu z korzyściami oraz 2) altruista i odbiorca muszą często ze sobą współdziałać i regularnie wymieniać się rolami. . Taki system sikora za siusia zamienia krótkoterminowy altruizm w długoterminową współpracę i trwa tak długo, jak długo korzyść jest zawsze większa niż koszt dla każdego aktora. Stabilny, wzajemny, altruistyczny związek jest równoznaczny z przyjaźnią. Aktorzy nie muszą być ze sobą powiązani, aby ten system działał, o ile jest zrównoważony. Wiele naczelnych, takich jak pawiany, tworzy koalicje męsko-męskie przeciwko samcom o wyższych rangach. Często pielęgnacja innej osoby będzie uważana za zaliczkę na przyszłą pomoc, jeśli zostanie zaatakowana przez osobę zagrażającą. Szympansy, które się nawzajem pielęgnują, częściej dzielą się mięsem z zabitych przez małpy colobus. Wzajemny altruizm działa tak długo, jak długo nie jest dostępny dla oszustów, którzy nie uczestniczą uczciwie. Tak więc wykrywanie oszustów ma kluczowe znaczenie, a umiejętność rozróżniania między przeciwsobnikami i nieodwzajemniaczami jest niezbędna. Oszustów należy unikać lub karać. Ponadto aktorzy muszą również być w stanie śledzić korzyści i koszty działań altruistów. Prawdopodobnie w tym miejscu pojawia się zwiększone funkcjonowanie poznawcze człowieka, ponieważ ludzie znacznie lepiej radzą sobie z długotrwałymi i złożonymi sieciami wzajemnego altruizmu niż szympansy i są od nich znacznie bardziej zależni. Społeczeństwa na dużą skalę oparte na zasobach rolnych, które pojawiły się po 10 Kya, są stosunkowo nowe. Przez większość ewolucji człowieka grupy były małe i utrzymywały się z polowań, zbieractwa i żerowania, ale potrzeba okablowania neuronowego do zarządzania wieloma sieciami wzajemnego altruizmu w przestrzeni i czasie była wyraźnie ważna nawet w małych grupach. Kiedy niektóre populacje ludzkie przyjęły siedzący tryb życia, oparty na uprawach, ludzie byli w stanie prosperować w społeczeństwach na dużą skalę, po części dzięki altruistycznym zachowaniom, które wybrano u ich przodków. Ludzie grupują się razem z wielopokoleniowymi rodzinami i krewnymi z większymi grupami rzędu 100-1 000 000 i jest to typowo pokojowy układ, ponieważ podstawą ludzkiej społeczności jest wzajemność (współpraca, koalicje i wymiana), co jest rzadkie u naczelnych nie będących ludźmi, chociaż większość naczelnych żyje w grupach. Istnieją różne powody, dla których naczelne ewoluują, by żyć w grupach. Zachowania w grupie mogą być napędzane potrzebą obrony zasobów (siła w liczebności) lub potrzebą ochrony przed drapieżnikami (bezpieczeństwo w liczebności). Ale grupy mogą ewoluować tylko wtedy, gdy korzyści dla jednostki przeważają nad kosztami. (Pamiętaj, że selekcja działa na jednostkę, a nie na grupę czy populację.) Korzyści dla jednostki obejmują dostęp do pożywienia (np. polowania grupowe), dostęp do partnerów, zmniejszenie liczby drapieżników i wspólną opiekę nad potomstwem. Koszty życia grupowego obejmują rywalizację o zasoby, rywalizację o partnerów, ryzyko drapieżników, ryzyko chorób i ryzyko pasożytów. W konsekwencji w grupach tworzą się hierarchie dominacji, w których niektóre osoby mają wyższą lub niższą rangę lub status niż inne osoby.
Początki Człowieków (58)
REWOLUCJA LUDZKA
Dowody na to, co uważamy za współczesną kulturę i zachowania - takie jak ozdabianie się ubraniami i biżuterią, tworzenie sztuki i wykonywanie rytuałów - nie pojawiają się w rzeczywistości, gdy pierwsi anatomicznie współcześni ludzie pojawiają się około 200 Kya. Biologia i zachowanie nie są bezpośrednio powiązane w ewolucji homininów, więc dowody na ludzkie zachowanie pojawiają się długo po ewolucji współczesnej anatomii człowieka. Chociaż ewolucja ludzkiej kultury i zachowań następowała stopniowo, zjawisko to określa się mianem rewolucji górnego paleolitu. Oczywiste jest, że homininy są ludźmi, zanim zostawią za sobą zaawansowane szczątki kulturowe, ale hipotetycznie możliwe jest cofnięcie człowieczeństwa w czasie, ponieważ ludzkie cechy skumulowały się przez miliony lat. Kultura nie jest unikalna dla ludzi, ale z pewnością jest przesadzona u ludzi i żadne inne zwierzę nie jest zależne od kultury tak jak my. Kultura jest zwykle definiowana jako ludzkie zachowanie i działania, które są regulowane przez zwyczaje i zasady społeczne, i jest utrwalany, ponieważ jest przekazywane z pokolenia na pokolenie z tradycją i nauką. Chociaż szympansy nie mają języka, samice w niektórych populacjach są w stanie podtrzymywać kulturę łowienia termitów, ponieważ matki uczą swoje potomstwo, jak żerować na owadach. Również tradycja rozłupywania orzechów przez szympansy kamieniami nie jest uniwersalna dla gatunku, co odpowiada różnicom między ludzkimi tradycjami kulturowymi. Istnieje kilka stwierdzeń dotyczących najwcześniejszej sztuki, ale daty są najlepsze dla kawałków czerwonej ochry z jaskini Blombos w RPA, w wieku około 79 Kya. Te małe fragmenty miękkiego czerwonego minerału są szlifowane, aby uzyskać płaską powierzchnię i celowo wygrawerowane "X" - bardzo podobne do czegoś, co można narysować na marginesach tej strony. Istnieją również przebite muszle, które byłyby używane jako koraliki do osobistego ozdabiania, które mogą datować się już na 100 Kya w Algierii, Izraelu i Afryce Południowej, ale daty nie są potwierdzone. Sztuka staje się coraz bardziej płodna i z biegiem czasu nabiera wartości muzealnej. Niektóre kojarzą się z neandertalczykami, jak proste wisiorki lub rowkowane i wypolerowane kości i zęby. Miejsca już w 400 Kya zachowały fragmenty czerwonych i czarnych pigmentów, które mogły być używane do ozdabiania ciał lub przedmiotów. Później, po 50 Kya, jasne jest, że te pigmenty zostały spalone i użyte jako farba. Jednak ludzie z górnego paleolitu pozostawili po sobie znacznie więcej niż tylko pigmenty, są flety, rzeźbione zwierzęta, rzeźbione kobiety i przepięknie malowane malowidła ścienne w jaskiniach, jak w słynnych jaskiniach Lascaux i Chauvet we Francji, które zawierają zmysły projektu, tekstury i koloru . Ogromna liczba statuetek tak zwanych "Wenus" znajduje się w całej Europie i Azji w górnym paleolicie. Wyrzeźbione z kamienia i kości słoniowej, te przenośne figurki to zawsze kobiety i charakteryzują się przesadnymi cechami anatomicznymi. Mają ogromne piersi, wystające brzuchy, szerokie biodra i zaznaczone złogi tłuszczu na udach, biodrach i pośladkach. Niewiele ma szczegóły twarzy, ale Brassempouy Lady, statuetka z kości słoniowej z Francji na 25 Kya, jest jednym wyjątkiem. Niektórzy archeolodzy uważają posągi za symbole płodności, prawdopodobnie odzwierciedlające boginię płodności. Być może najbardziej rozpoznawalną "Wenus" jest rzeźbiona kobieta Willendorf z Austrii w wieku około 23 Kya. Najstarsze malowidła naskalne w Afryce znajdują się w Namibii i datowane są na około 27 Kya, ale to tylko te, które do tej pory były rygorystycznie datowane. Niektóre afrykańskie sztuki naskalne mogą datować się na ponad 70 Kya, ale zazwyczaj są one przyćmione w literaturze przez europejską sztukę jaskiniową ze względu na lepszą ochronę, a przede wszystkim w Europie jest więcej zajętych jaskiń. Względna obfitość sztuki i artefaktów w Europie w porównaniu z Afryką w górnym paleolicie wynika z błędów konserwatorskich. Ludzie z górnego paleolitu pozostawili również dowody na swoje wyszukane zdobienia ciała. Niektóre z najstarszych, dobrze datowanych biżuterii to niektóre koraliki ze strusich skorupek z otworami do łączenia ich ze sobą, które zostały odkryte w Turcji między 45 a 40 Kya. Mniej więcej w tym samym czasie podobne artefakty znaleziono w Kenii, Libanie i Bułgarii. Kultura Châtelperronian znana jest z używania zębów - często od mięsożerców, takich jak lisy, niedźwiedzie, wilki i hieny - w biżuterii i ozdabianiu ciała. Nie zachowała się żadna odzież z górnego paleolitu, ale filogenetyka molekularna wszy zawiera pewne wskazówki. Wszy głowowe i wszy ciała były kiedyś tym samym gatunkiem. Wszy głowowe żywią się skórą głowy, a wszy żywią się ciałem, ale żyją w ubraniu. Szacuje się, że pochodzenie wszy ciała lub czas ich oddzielenia od wszy głowowych miało miejsce między 72 a 42 Kya, co oznacza, że prawdopodobnie w tym czasie przyjęto odzież (siedlisko dla wszy). Ponieważ DNA wszy afrykańskich ciała jest bardziej zmienne niż DNA wszy ciała na całym świecie, jest zgodne z ekspansją wszy ciała i ich odzianych nosicieli z Afryki. Język, jak omówiono powyżej, a także religia są cechami tradycyjnie wrzucanymi do ludzkiej rewolucji, ponieważ wiążą się z myśleniem symbolicznym. Dowody na wczesną religię i rytuały pochodzą głównie z pochówków. Czasami można stwierdzić, za pomocą kontrolowanego wykopu, czy szkielet został celowo zakopany, czy nie. Istnieje różnica w osadzie związanym z porównywanym ciałem do tego tuż nad i pod nim. Również celowe przetwory grobowe szkielety są znacznie lepsze niż pozostawione na powierzchni, aby padlinożercy, pogoda i procesy środowiskowe rozproszyły lub zniszczyły kości. Celowy pochówek niekoniecznie musi być utożsamiany z rytuałem lub religią; może to być po prostu sposób na prowadzenie domu. Dowód na celowość pojawia się, gdy szkielet znajduje się schowany w pozycji płodowej, ale krytycy twierdzą, że za to może odpowiadać wygoda (mniejszy grób), a nie rytuał. Celowość pochówków ludzkich nie jest przedmiotem dyskusji. Występują na wolnym powietrzu i w jaskiniach i obejmują koraliki, narzędzia, ochrę, węgiel drzewny i inne przedmioty. Możliwe, a dla niektórych jest to niewątpliwe, że neandertalczycy celowo chowali także swoich zmarłych. Istnieją dowody potwierdzające pochówek neandertalczyków w miejscach takich jak Shanidar w Iraku (60 Kya), gdzie pyłek kwiatowy został wydobyty ze szczątków szkieletu neandertalczyka. Pyłek mógł wskazywać, że neandertalczycy odprawiali rytuały i czcili życie i śmierć. Krytycy twierdzą, że pyłek mógł nagromadzić się przypadkowo i niekoniecznie jest wynikiem rytuału lub wiary w życie pozagrobowe. Poza tym, sądząc po urazie szkieletu Shanidara, osoba ta wyraźnie wiodła trudne życie. Ciężki cios w lewą stronę czaszki najprawdopodobniej częściowo go oślepił, a jego ramię częściowo amputowano i wyleczono. Poczucie długotrwałej straty mogło zacząć się u ludzi już w 160 Kya, ponieważ z wypolerowanych czaszek znalezionych w miejscu Herto w Etiopii jasno wynika, że ktoś je trzymał i zajmował się nimi dawno po śmierci.
Początki Człowieków (57)
JĘZYK
Język jest tak integralną częścią bycia człowiekiem, że nawet się go nie uczy. Zamiast tego język rozwija się naturalnie jak narząd lub to, co Steven Pinker nazywa "instynktem językowym". W wieku pięciu lat ludzie znają wszystkie zasady języka. Noam Chomsky nazwał naszą automatyczną skłonność do uczenia się i opanowywania skomplikowanych, a czasem nielogicznych reguł gramatyki, naszym "urządzeniem przyswajania języka", a jego teorię wzmocniło odkrycie "gramatyki uniwersalnej", która jest powszechną podstawą dla wszystkich ludzkich języków. Bardzo trudno jest znaleźć w zapisie kopalnym bezpośrednie dowody na ewolucję mowy i języka człowieka. Żadna z miękkich części anatomicznych biorących udział w mowie, takich jak język, krtań (skrzynka głosowa) i podniebienie miękkie, nie jest zachowana, a części kostne, które ulegają fosylizacji, nie wykazują zbyt użytecznej anatomii. Nadal nie ma jednoznacznych dowodów na to, że jakikolwiek hominin inny niż współcześni ludzie miał język lub mógł nawet mówić tak jak my. Uważano, że mała, pływająca kość w kształcie podkowy w szyi, zwana gnykową, zawiera pewne wskazówki, a ponieważ zachowała się ona u Neandertalczyka Kebara (od 60 Kya w Izraelu), nadzieje były duże. Stwierdzono jednak, że anatomia kości, chociaż podobna do ludzkiej, a nie do szympansa, jest identyczna z kością świni, więc nie jest diagnostyczna dla zdolności językowych. Chociaż, biorąc pod uwagę długość szyi oraz podstawę twarzy i szczęki, trakt głosowy neandertalczyków wydaje się mieć proporcje małpy. Małpy człekokształtne i ludzkie niemowlęta mają wysoką krtań i mogą jednocześnie ssać i oddychać. Ale gdy ludzie rozwijają się w przeszłości niemowlęctwa, ich krtań obniża się i to pomaga nam wydawać wiele dźwięków samogłosek w gardle, ale uniemożliwia nam jednoczesne oddychanie i jedzenie. Ponieważ określone obszary mózgu są powiązane z mową i językiem, może istnieć możliwość zebrania dowodów na język z endocastów mózgu. Na przykład, czapka konkretnego H. erectus cranium SM 3 (z Sambungmacan w Indonezji) ma wyraźną czapkę Broca, guzek na lewym płacie czołowym odpowiadający obszarowi Broca, który jest obszarem anatomicznym związanym z produkcją mowy i przetwarzaniem języka. Ale czapki Broca można znaleźć na odlewach z szympansów, więc nie są niezawodnymi wskaźnikami języka. Co więcej, mózg jest owinięty warstwą opon mózgowo-rdzeniowych, która składa się z warstw zwanych pia mater, pajęczynówką i oponą twardą, która zaciemnia wrażenie mózgowe w przypadku odlewu, co sprawia, że interpretacja guzków w mózgu jest jeszcze trudniejszym zadaniem. Skomplikowana koordynacja i kontrola mięśni tułowia są niezbędne do kontrolowania precyzyjnie dostrojonych wzorców oddychania podczas mowy ludzkiej. Możliwe jest więc powiązanie rozmiaru rdzenia kręgowego z ilością nerwów potrzebnych do monitorowania oddychania i mowy. Rozmiar otworu w kręgu, otwór kręgowy, jest mały u H. erectus (w oparciu o chłopca Nariokotome), co sugeruje, że jego rdzeń kręgowy również był mały. Tak więc u H. erectus prawdopodobnie nie było tak silnego osłabienia klatki piersiowej, jak u współczesnych ludzi i jest mało prawdopodobne, by chłopiec Nariokotome mógł mówić tak jak my. Ale to nie wyklucza możliwości, że miał szczątkowy język. Ponieważ dowody anatomiczne są niejednoznaczne, nie możemy wykluczyć, że jakiś konkretny hominin ma język. Jednak naukowcy mają tradycyjnie skorelowano symbole, sztukę oraz ludzką kulturę i technologię z obecnością języka i nic z tego nie pojawia się aż do 100 Kya. Jednak polowanie kooperacyjne, z dowodami na wypędzanie stad z klifów w środkowym plejstocenie, może być wcześniejszą innowacją, którą można powiązać z językiem. Złożoność społeczna (a tym samym funkcjonowanie poznawcze, które jest wymagane dla języka) związana ze wspólnym polowaniem istnieje na znacznie większą skalę niż ta, którą osiągają psy i koty polujące w grupach. Genetyka mowy i języka zaczyna się rozwijać. Mutacje w genie FOXP2 powodują, że osoba ma problemy z kontrolą motoryczną ust i mięśni twarzy, przez co wymowa słów jest trudna. Osoby z mutacjami w genie mają również braki w pewnych aspektach gramatyki i poznania. Po zidentyfikowaniu FOXP2 naukowcy szukali jego funkcji u normalnych ludzi i innych zwierząt. FOXP2 jest genem regulatorowym, występującym w podobnych formach przez wszystkie kręgowce, który zarządza czynnościami innych genów, z których niektóre są zaangażowane w język, a inne w ogóle nie są zaangażowane w język (co komplikuje hipotezy dotyczące jego selekcji wyłącznie na podstawie języka) . Ludzka wersja genu różni się od myszy tylko trzema aminokwasami, a od szympansa tylko dwoma, ale możliwe, że te dwie zmiany miały znaczenie funkcjonalne dla pochodzenia języka. Zegary molekularne ludzkiego FOXP2 wskazują na bardzo niedawne pochodzenie w ciągu ostatnich 200 000, ale ponieważ jest to gen regulatorowy, jest wysoce nieprawdopodobne, aby był jedynym genem zaangażowanym w ewolucję zdolności językowych. FOXP2 przypomina nam, że ewolucja to "pochodzenie z modyfikacją", jak powiedział Darwin, ponieważ coś tak istotnego dla uczynienia ludzi "człowiekami" prawdopodobnie wyewoluowało z prymitywnych genów, które wcale nie są unikalne dla ludzi. Selekcja musiała działać bardzo silnie na język, tak jak działała na dwunożność. Hipotezy dotyczące jej najwcześniejszych zalet obejmują zdolność do skuteczniejszego polowania poprzez wymianę informacji o środowisku fizycznym i ekologicznym. Robin Dunbar sugeruje, że język ewoluował w celu wymiany informacji o środowisku społecznym, a właściwie plotkowania. Komunikacja społeczna nie jest trywialną sprawą, ponieważ informacje o tym, komu można ufać - to znaczy kto uczciwie gra we wzajemność - mogą być udostępniane całej społeczności (zobacz sekcję "Altruizm i ludzka kolonia" w tym rozdziale). Dzięki językowi jednostka nie musiałaby śledzić wszystkich złożonych relacji i sieci społecznych i politycznych w swojej społeczności. Plotkowanie ułatwiało przekazywanie i zapamiętywanie tych informacji. Język w tym sensie pozwalał ludziom żyć w społeczności w dużych grupach i zwiększał znaczenie reputacji jednostki, jeśli inni mogli udostępniać informacje o niej lub jej opinii publicznej. Język mógł być również, jak sugeruje Geoffrey Miller, narzędziem uwodzenia preferowanym przez dobór płciowy. Naczelne w dużym stopniu polegają zarówno na języku głosowym, jak i niegłosowym, a u naczelnych istnieją podstawy złożonej komunikacji międzyludzkiej. Jako całość grupa wykorzystuje zwiększoną liczbę mięśni twarzy do przekazywania emocji. Gibony białoręczne używają specjalnych pieśni do ostrzegania innych, że drapieżnik jest niebezpiecznie blisko. Szympansy są świadome znaczenia komunikacji mimicznej i zaobserwowano, że dosłownie wycierają z twarzy uśmiechy strachu w obecności agresywnego członka grupy, aby wyglądać na nieustraszonego. Szympansy trzymane w niewoli w naturalny sposób wytwarzają specyficzne wołania o określone produkty spożywcze, takie jak banany, a dzikie szympansy używają specyficznych odgłosów dla węży i zagrażających nieznajomym, a także mają nawoływania do polowań.
Początki Człowieków (56)
REPRODUKCJA
Dla niektórych, takich jak Owen Lovejoy, w reprodukcję zaangażowane są naprawdę odrębne cechy ludzkie. Kobiety są podatne na kopulację, nawet gdy nie mają owulacji, pary łączą się w pary prywatnie i często łączą się w pary tylko z jednym partnerem przez długi czas, a nawet przez całe życie. Jest to uderzający kontrast z zachowaniami reprodukcyjnymi pawianów i szympansów, które mają wiele podobieństw z ludźmi pod względem biologii, społeczeństwa i inteligencji. Samice pawianów i szympansów zwykle łączą się w pary tylko wtedy, gdy są w rui, co jest częścią ich cyklu owulacyjnego, kiedy ich krocze odbytowo-płciowe nabiera jasnoróżowego obrzęku, sygnalizując nadejście ich płodności. Często obfitość pożywienia powoduje, że samice z grupy wchodzą w ruję, a samce osiągają podwyższony poziom konkurencji wokół tych samic. Ludzkie samice poczęły owulację, która jest ukryta zarówno przed samcami, jak i przed nimi samymi. Ta sytuacja otwiera ich na nieustanne możliwości łączenia się w pary i może wymagać od samców znalezienia alternatywnych sposobów wykrywania owulacji żeńskiej, takich jak na przykład inne wskazówki wizualne, a nawet węchowe (zapachowe). Dodatkowo ukryta owulacja może wprowadzić samców w stan ciągłej rywalizacji. Oczywiście w światach pawianów i szympansów jest znacznie mniej prywatności, ale mogą się kojarzyć potajemnie, jeśli naruszone zostaną kody statusu i hierarchii. Ponadto, w przypadku większości naczelnych, częściej lepiej kojarzyć się z więcej niż jednym partnerem, nie tylko w jednym cyklu godowym, ale przez całe życie. Większość ludzi stosuje strategię monogamiczną, to znaczy tworzą trwałe więzi par (seryjne lub na całe życie), ale wielu ludzi jest poligamicznych, tworząc zamiast tego długotrwałe relacje między jednym mężczyzną a wieloma kobietami. W obu przypadkach konkurencja między ludźmi jest znacznie mniejsza, a także niezwykle wysoki poziom zaangażowania rodziców w mężczyzn. Ze względu na ich wydłużony okres wzrostu i dojrzewania, głównie z powodu niewiarygodnej ilości czasu potrzebnego do wyhodowania dużego ludzkiego mózgu, ludzkie niemowlęta są altrialne, co oznacza, że po urodzeniu są wrażliwe i wysoce zależne od rodziców. Odwrotnym warunkiem jest bycie przedwcześnie rozwiniętym, jak nowonarodzone konie, które mogą zacząć biegać zaraz po dotknięciu ziemi, ponieważ osiągają większy wzrost w macicy. Ludzie wyraźnie mają własną strategię reprodukcyjną, która różni się nawet od ich najbliższych krewnych, a my mamy do tego wyjątkową anatomię. H. sapiens wykazują niski poziom dymorfizmu płciowego w wielkości ciała, sile, fizjologii i są najmniej dymorficzne spośród wszystkich małp człekokształtnych, ale nie jesteśmy asmonomorficznymi amonogamicznymi gibonami. Mężczyźni są więksi i silniejsi i mają wyższe tempo metabolizmu niż kobiety. Mają wyższy wskaźnik śmiertelności nieletnich i osiągają dojrzałość płciową później niż samice. Rozmieszczenie i obfitość włosów i tłuszczu różnicują płcie, podobnie jak nieco duże i rzucające się w oczy narządy płciowe. Ludzie dzielą się między szympansy (duże) i goryle (małe) pod względem wielkości jąder dla masy ciała. Innymi słowy, ludzie mają jądra pomiędzy wysoce konkurencyjnymi (szympansy) a zasadniczo wolnymi od konkurencji (goryle) jądrami. Samce z gatunku naczelnych, w których samice kojarzą się regularnie z więcej niż jednym samcem, mają zwykle wymyślne penisy, które są albo silnie stymulujące, albo podobne do szczeciny do bezpośredniej rywalizacji plemników w pochwie. Gatunki, w których samice łączą się w pary tylko z jednym samcem podczas cyklu rujowego, mają znacznie mniej skomplikowaną anatomię i przypominają ludzką formę. Jednak ludzki penis jest znacznie większy i nie ma baculum (kości prącia), co niektórzy sugerują, że jest wynikiem doboru płciowego przez kobiety, zwłaszcza że dwunożność sprawiła, że męskie genitalia są znacznie bardziej widoczne. Dwunożność mogła również pomóc ukształtować anatomię seksualną ludzkich kobiet. Piersi są większe niż oczekiwano, gdy nie karmią piersią, a rozkład tłuszczu na biodrach, udach i pośladkach obniża wskaźnik talia-biodra (WHR). Oba są prawdopodobnie sygnałami płodności i wskaźnikami sprawności. Wartość WHR wynosząca 0,7 jest niemal krzyżowo uważana za "idealną" kobiecą sylwetkę, niezależnie od tego, czy kobieta jest szczupła, czy ciężka. Kobiety o klasycznej sylwetce klepsydry mają optymalny poziom estrogenu, który jest skorelowany z płodnością. Więc chociaż żaden samiec nie mierzy i nie oblicza WHR potencjalnych partnerów, są oni w stanie wychwycić sygnały z ciała kobiety, które są powiązane z ewolucyjną sprawnością. Australopity miały wysoki stopień dymorfizmu płciowego w ciele i wielkości psów, który był większy niż u ludzi, więc strategia rozrodu człowieka nie ewoluowała u australopitów. Wczesne homininy były prawdopodobnie poligyniczne w swoich zachowaniach godowych, jak współczesne szympansy, ale do czasu pojawienia się H. erectus strategie uległy zmianie. Dowody kopalne wskazują, że dymorfizm płciowy znacznie zmniejszył się u H. erectus. Ponadto gatunek urósł w rozmiar ciała, ale zachował niewielką szerokość bioder (na równi z australopitami) ze względu na sprawność dwunożną oraz adaptację klimatu w proporcjach ciała. Wzrostowi wielkości mózgu musi towarzyszyć wzrost wielkości ciała, więc H. erectus nie mógł zmieścić się w mózgu i czaszce niemowlęcia tego samego rozmiaru, jak oczekiwano dla jego wielkości ciała poprzez wąskie biodra. Oznacza to, że H. erectus mógł mieć skróconą ciążę (tj. okres rozwoju płodu w macicy), aby fizycznie móc urodzić dzieci z większym mózgiem przez stosunkowo mały kanał rodny. Wcześniejszy poród skutkuje bardziej bezradnym, słabiej rozwiniętym, altrialnym niemowlęciem. Jest więc prawdopodobne, że wraz z H. erectus zaczął ewoluować wyższy poziom inwestycji rodzicielskich, zwłaszcza ze strony ojca (inwestycja ojcowska). Selekcja sprzyjałaby takiej zmianie, gdyby sprzyjała wzrostowi i rozwojowi mózgu, zwłaszcza jeśli dobór działał silnie na wzrost wielkości mózgu u gatunku. Inne ważne zmiany są ze sobą skorelowane, takie jak włączenie mięsa do diety, dzielenie się pożywieniem i tworzenie baz domowych, w których samce zaopatrywały samice i potomstwo oraz gdzie samice lokalizowały swój duży wkład w dietę rodzinną dzięki żerowaniu i zbieraniu (podział pracy ze względu na płeć). .Gdy duże inwestycje ze strony ojca w potomstwo i tworzenie więzi parowych stałyby się udanymi adaptacjami, samice wybrałyby również kojarzenie się i wiązanie z samcami, które nie tylko zapewniały pożywienie i ochronę, ale posiadały niezawodne i ojcowskie cechy, takie jak życzliwość, hojność i wiarygodność . Mniej czasu spędza się na żerowaniu, gdy jakość jedzenia rośnie, więc dzięki dodaniu do diety mięsa i innych wysokiej jakości pokarmów, prawdopodobnie więcej wolnego czasu spędzano w bazie domowej. Kiedy to miało miejsce, cechy intelektualne, takie jak wyszukany język, śpiew, muzyka, dowcip, a nawet taniec stałyby się bardzo ważne w doborze partnera. Nie ma dowodów na to, że język ewoluował już w czasach H. erectus, ale nie można wykluczyć śpiewu, tańca i humoru we wczesnym plejstocenie.
Klimakterium
Menopauza, czyli przerwanie reprodukcji, to ewolucyjna zagadka. W jaki sposób dobór naturalny mógłby go faworyzować, skoro wyraźnie preferuje zwiększoną reprodukcję? Kobiety doświadczają menopauzy około pięćdziesiątego roku życia, kiedy ich ograniczona podaż komórek jajowych jest wyczerpana, a procesy rozrodcze spowalniają i ostatecznie ustają. Niektórzy, w tym Kristen Hawkes i jej współpracownicy, uważają, że menopauza może być cechą adaptacyjną. Kobiety, które przestają rozmnażać się w okresie menopauzy, zaczynają pomagać swoim aktywnym reprodukcyjnie córkom w utrzymaniu wnuków. Podczas gdy jej córka laktuje (jest to energetycznie kosztowny proces), babcia pomaga dofinansowywać dietę dzieci niekarmiących piersią, zbierając dodatkową żywność. W ten sposób babcie pomagają swoim córkom mieć więcej dzieci, a także zwiększają ich przeżywalność. Przypuszczalnie materiał genetyczny babci, który zawiera geny umożliwiające długie przeżycie po menopauzie, jest przekazywany jej lepiej przeżywającym dzieciom i wnukom. Inne wyjaśnienie menopauzy, przedstawione przez Jamesa Wooda i współpracowników, nie uważa jej za adaptację, pomimo korzyści płynących z opieki babci. Zgodnie z hipotezą Wooda, menopauza i życie po menopauzie same w sobie nie są korzystne - raczej ewoluowały z powodu antagonistycznej plejotropii, procesu, w którym geny, które mają szkodliwe skutki w późniejszym życiu, mogą być aktywnie selekcjonowane, jeśli mają korzystny wpływ we wcześniejszym okresie życia. U młodych kobiet proces znany jako atrezja pęcherzyków pomaga zmaksymalizować płodność, ale także powoduje przyspieszenie układu rozrodczego poprzez ograniczoną podaż jaj. Menopauza jest zatem kompromisem, a nie adaptacją, ponieważ kobiety rezygnują z możliwości rozmnażania się w późniejszym życiu, aby we wczesnym okresie życia doświadczyć wysokiego poziomu płodności. Być może nie powinniśmy pytać, dlaczego menopauza wyewoluowała, ale zamiast tego zbadać, kiedy kobiety zaczęły żyć poza okresem rozrodczym i dlaczego kobiety, w porównaniu z mężczyznami, mają przede wszystkim ograniczoną produkcję gamet.
Początki Człowieków (55)
OGIEŃ
Manipulacja ogniem doprowadziła do dramatycznych zmian w zachowaniu homininów. Ogień oferuje ciepło i umożliwił homininom rozszerzenie ich siedliska na zimniejsze regiony (albo na wyższych wysokościach, albo na wyższych szerokościach geograficznych). Ogień zapewnia światło w nocy lub w jaskiniach; może służyć jako ochrona przed drapieżnikami; może być używany podczas polowań do kontrolowania ruchów zwierząt; przydaje się do robienia narzędzi z drewna i niektórych rodzajów kamieni, które najlepiej utwardzić pod wpływem ciepła; oferuje również sposób na detoksykację i zmiękczenie żywności (zarówno roślinnej, jak i zwierzęcej) poprzez gotowanie. Archeologiczne dowody pożaru obejmują nagromadzenie popiołu i spaloną ziemię, skały, kości, drewno, węgiel drzewny lub inne artefakty. Przebarwienia oraz zmiany chemiczne i strukturalne tych materiałów wskazują na spalanie, a zmiany atomowe zachodzące po przegrzaniu obiektów można śledzić za pomocą termoluminescencji. Najwcześniejsze dowody kontrolowanego pożaru znajdują się w Gesher Benot Ya′aqov, miejscu w Izraelu, datowanym na 790 Kya. Posiada narzędzia aszelskie, spalone artefakty krzemienne, fragmenty węgla drzewnego, spalone drewno, owoce i ziarna. Stanowisko H. erectus w Zhoukoudian w Chinach ma również zachowane palenisko datowane na około 500 kya, podobnie jak miejsce zabicia H. erectus/archaiczne w Terra Amata we Francji, na około 380 kya. Krótkoterminowe ślady ognisk (gleby w kształcie misy) zostały odkryte na znacznie wcześniejszych stanowiskach, takich jak Koobi Fora (1,6 miliona lat temu) i Chesowanja (1,4 miliona lat temu) w Kenii. Istnieją również spalone kości w jaskini Swartkrans w Afryce Południowej od około 1,5 miliona lat temu. Ale te bardziej starożytne miejsca nie zostały jeszcze potwierdzone jako miejsca z kontrolowanym ogniem. Po ustaleniu, że rzeczywiście istnieją dowody na istnienie ognia, trudno jest odróżnić jego naturalne istnienie od ludzkiej kontroli nad nim. Wysokie temperatury spowodowane koncentracją ognia w palenisku przewyższają te występujące w przypadku naturalnych pożarów, które rozprzestrzeniają się po krajobrazie. Jak dotąd okazało się niemożliwe do ustalenia, czy ludzie oportunistycznie kontrolowali ogień (pobrany z naturalnie występującego źródła), czy też celowo go wywołali. To ostatnie oznacza poziom wyrafinowania, który jest "ludzki", podczas gdy pierwszy nie. Chociaż oportunistyczne użycie ognia jest dużym krokiem przed małpami, bez względu na to, w jaki sposób go uzyskano. Kontrolowany ogień i korzystanie z palenisk w bazach domowych i miejscach zamieszkania wyraźnie stały się popularne do czasu, gdy archaiki stały się centralnym punktem ewolucji homininów.
Początki Człowieków (54)
PADLINOŻERNOŚĆ I POLOWANIE
Ludzie mają wspólnego roślinożernego przodka z żywymi małpami, a wczesne homininy były głównie wegetarianami, którzy jedli owoce, orzechy, bulwy (korzenie), a także owady, takie jak termity. Podobnie jak inne ssaki roślinożerne, w tym małpy i małpy człekokształtne, ludzie nie potrafią syntetyzować witaminy C - niepotrzebna umiejętność przy diecie złożonej z materii roślinnej bogatej w witaminę C. Jednak drastyczna zmiana wielkości mózgu i ciała homininów w okolicach 2 milionów lat temu wiąże się ze zmianą diety w kierunku mięsożerności. Zbieranie i polowanie podczas gorących godzin dziennych na sawannie wschodnioafrykańskiej jest ważnym elementem ewolucji zestawu postaci, które odróżniały wczesne Homo od australopitów i parantropów i ostatecznie doprowadziły do powstania ludzi. Chociaż jedzenie mięsa zajmuje ważne miejsce w naszej ewolucyjnej historii, nie pojawiło się w naszej linii jako jedynej. Głównie owocożerne szympansy w Gombe w Tanzanii znane są z tego, że mają zwyczaj zabijania i zjadania czerwonych małp colobus do tego stopnia, że znacznie zmniejszają populację małp w regionie. Znane są nawet z zabijania małych byków i dzikich świń. Szympansy używają również narzędzi do polowania na mięso. Jill Pruetz i Paco Bertolani obserwowali samce, samice, a nawet młode szympansy w Senegalu wytwarzające włócznie z gałęzi (podobnie jak samice wytwarzają różdżki do połowu termitów) i używając ich do zabijania małych zwierzaków (nocne strepsiryny, które śpią w ciągu dnia). Wbijali włócznie w puste drzewa, w których gnieździły się młode , aby je unieruchomić przed włamaniem się do drzewa, aby je złapać i zjeść. Trudno sobie wyobrazić, że naszym przodkom homininom brakowało takiego wyrafinowania łowieckiego, ale niestety włócznie, takie jak te wykonane przez szympansy, nie zachowały się w zapisie kopalnym, abyśmy mogli się o tym przekonać. Aby śledzić ewolucję ludzkiego jedzenia mięsa poprzez padlinożerstwo i polowanie, musimy śledzić zapis narzędzi kamiennych i pożeranych kości zwierząt. Narzędzia kamienne znajdują się już w wieku 2,6 miliona lat temu, a wkrótce potem, około 2,5 miliona lat temu, najwcześniejsze skamieniałości bydła po cięciu pochodzą z Bouri w Etiopii. Chociaż te daty są zwykle przytaczane jako najwcześniejsze znaczące włączenie mięsa do diety homininów, żadne miejsce nie ma zarówno naciętych kości, jak i narzędzi kamiennych. Łóżka w wąwozie Olduvai (datowane już na około 2,5 Mya) zapewniają bezpośredni związek między narzędziami a wyrżniętymi kośćmi. Pat Shipman i Rick Potts badali ślady nacięć i ślady zębów mięsożerców na zbiorowisku fauny Olduvai i zasugerowali, że homininy zjadały mięso. Ślady nacięć na kościach wskazują na rzeź i pod mikroskopem można je odróżnić od śladów zębów mięsożernych. Płatek kamienia, bez względu na to, jak drobnoziarnisty jest materiał, nigdy nie będzie miał idealnie prostej krawędzi, tak że podczas przeciągania po kości podczas użytkowania, jego postrzępiona krawędź pozostawia wiele cienkich równoległych linii. Wręcz przeciwnie, zęby mięsożerców pozostawiają na kościach pojedynczy, zaokrąglony rowek. W Olduvai ślady nacięć nakładały się na ślady zębów i vice versa, co oznaczało, że albo homininy były głównymi myśliwymi/ padlinożercami, a mięsożercy wtórnie padli ofiarą z miejsc uśmiercania, albo że homininy padały ofiarą zabitych zwierząt mięsożernych. Drugi scenariusz, z homininami jako padlinożercami, ma zwykle miejsce w najwcześniejszych miejscach. Szpik wewnątrz kości jest szczególnie pożywny, a blizny po rozbijaniu kości kamiennymi narzędziami można również odróżnić od śladów po miażdżących kości zębach mięsożercy. Wobec braku śladów zębów mięsożernych do polowania w porównaniu do dowodów na padlinożerstwo, istnieją inne sposoby na ustalenie jedzenia mięsa. Ślady po cięciu narzędziami kamiennymi zlokalizowane w okolicach stawów szkieletowych są oznakami rzezi (np. w stawach, gdzie więzadła i ścięgna łączą kości). W zależności od tego, czy stanowisko archeologiczne jest miejscem zabicia (gdzie zwierzę jest zabijane i przewożone "do domu"), czy miejscem życia (lub "żywą podłogą"), brak lub obecność kości związanych z mięsnymi i smacznymi częściami zwierząt wskaż również, czy homininy miały pierwsze ślady na tuszy, czy też padły po drapieżniku. Nawet jeśli nie ma śladów nacięć, czasami możliwe jest zidentyfikowanie skamieniałego drapieżnika, ponieważ każdy gatunek drapieżnika ma szczególny sposób pożerania zwierzęcia. Niektórzy jedzą stopy i żebra i zostawiają resztę kości, ale niektórzy jedzą wszystko, nawet kręgi, pozostawiając tylko czaszkę i zęby. Bob Brain był jednym z pierwszych, który udokumentował różne preferencje żywieniowe drapieżników, ponieważ karmił je tuszami pawianów, a następnie analizował pozostałe części. Wybór homininów, którzy otrzymali mięso, musiał być bardzo wysoki ponieważ zbieranie z zabitych mięsożerców byłoby niezwykle niebezpieczne. Homininy mogły opracować przebiegłe metody skradania się w kierunku padliny, ale także mogły skoordynować grupy, aby odpędzić drapieżniki z ich pożywienia. Po zdobyciu umiejętności odpędzania drapieżnika przed zabójstwem, polowanie i zabijanie własnej ofiary przez homininy nie wydaje się wielkim skokiem behawioralnym. Ponieważ szympansy nie pozyskują mięsa poprzez padlinożerstwo, niekoniecznie zakładalibyśmy, że homininy też to zrobiły, gdyby nie liczne dowody z miejsc takich jak wąwóz Olduvai oraz nakładanie się śladów nacięć i zębów. Ewolucja jedzenia mięsa, a następnie zachowań drapieżnych jest prawdopodobnie źródłem wielu współczesnych przedsięwzięć sportowych i lekkoatletycznych. Biegi wytrzymałościowe i rzucanie przez ramię bez wątpienia pomogły wczesnym homininom doskonalić swoje umiejętności łowieckie. Geny takie jak ACE, chociaż nie występują we wszystkich populacjach, pomóż wyjaśnić, w jaki sposób ludzie mogą pracować ciężej przy mniejszym zużyciu paliwa i zwiększyć swoją wytrzymałość. Rzucanie miałoby ogromne znaczenie, ponieważ pozwala na działanie na odległość. A William Calvin zasugerował nawet, że ze względu na złożoną kontrolę neuronalną zaangażowaną w dokładne rzucanie ramieniem, jest to powiązane ewolucyjnie ze wzrostem wielkości mózgu i językiem. Spożywanie mięsa mogło być zdecydowanie korzystne i być może konieczne do wyhodowania dużego mózgu. Leslie Aiello postulował, że aby zrównoważyć energetyczny koszt wzrostu i utrzymania dużego mózgu, homininy wyewoluowały krótsze jelita (tj. selekcja faworyzowała homininy z krótszymi segmentami jelit). W porównaniu z małpami człekokształtnymi ludzie mają stosunkowo mniejsze wnętrzności, co jest cechą wspólną z mięsożercami. Ale materiał jelitowy jest prawie tak samo kosztowny energetycznie jak mózg i być może organizm jest narażony na rozwój "droższej tkanki" w mózgu, a mniej w jelitach, wzmacniając w ten sposób zmianę diety. Jedzenie mięsa i powiązane strategie oddzieliły Paranthropus od linii Homo. Podczas gdy Paranthropus żywił się materiałami roślinnymi i termitami, wczesne Homo zmieniło swoją dietę na mięso i szpik kostny, co być może wskazuje na ich specjację sympatryczną. Mniej więcej w czasie, gdy widzimy obfitość danych archeologicznych na temat jedzenia mięsa, następuje zmiana w wielkości ciała homininów i wielkości mózgu. Rosnący mózg potrzebuje więcej energii do utrzymania, a włączenie mięsa do diety mogło przyczynić się do większego mózgu i zwiększonej inteligencji (potrzebnej do umiejętności łowieckich) podczas karmienia. Można poczynić kilka prognoz dotyczących gatunku, który staje się drapieżnikiem. Opierając się na wiedzy z naszego obecnego zapisu kopalnego, H. erectus pasuje do wielu z tych przewidywań. Mianowicie H. erectus wykazuje wzrost wielkości ciała, zwiększenie zasięgu geograficznego, poważną zmianę technologiczną (z narzędzi Olduwańskich na narzędzia aszelskie) oraz wzrost towarzyskości. Ostatni punkt jest dyskusyjny, ponieważ dowody są pośrednie, ale niektórzy wskazują na ewolucję polowań grupowych jako wsparcia.
Początki Człowieków (53)
DIETA
Paleoantropolodzy interesują się ewolucją diety, ponieważ jest ona tak silnie skorelowana z innymi zmiennymi (takimi jak rozmiar mózgu, rozmiar ciała, inteligencja, poziom aktywności, zasięg geograficzny, kształt zębów, kształt czaszki itp.) i, co nie bez znaczenia, ponieważ spożycie pokarmu jest podstawowym środkiem przetrwania. Dieta również znacznie się zmieniła podczas ewolucji człowieka. Jak już wspomnieliśmy, narzędzia i zęby dostarczają wskazówek do diety homininów. Istnieje jednak kilka innych przydatnych metod rekonstrukcji diety homininów. W diecie obowiązują zasady ekologiczne i energetyczne. Energia słoneczna jest włączana do materii roślinnej, która jest następnie włączana do mięsa roślinożerców i ostatecznie pobierana przez mięsożerców. Na każdym poziomie piramidy troficznej (tj. uporządkowanie grup zwierząt według diety, z drapieżnikami na szczycie piramidy) spadek energii dostępnej na jednostkę biomasy. Z reguły, naczelne o dużych rozmiarach ciała mają tendencję do zjadania roślinności ze względu na jej obfitość w stosunku do nektaru i owadów, które są wystarczające do odżywiania gatunków o małych ciałach. Dosłownie jesteśmy tym, co jemy. Pierwiastki śladowe w skamieniałych zębach i kościach mówią o poziomie troficznym osobnika. Na przykład, metaliczny stront (Sr), który jest zwykle badany w odniesieniu do wapnia jako stosunek Sr/Ca, jest pobierany przez rośliny przez wody gruntowe, ale jest pobierany tylko przez zwierzęta, które jedzą rośliny. Niski poziom Sr wskazuje na mięsożerność, a wysoki na roślinożerność. Ten sam rodzaj informacji troficznej można również uzyskać z poziomu baru, magnezu i cynku. Homininy w Swartkrans mają wszystkożerne (pośrednie) poziomy Sr, co przeczy powszechnemu założeniu, że australopity i Paranthropus były czysto roślinożerne. Stabilne izotopy z zębów i kości również opowiadają o diecie. Izotopy węgla są szczególnie pouczające i opierają się na charakterystyce dwóch różnych ścieżek wiązania dwutlenku węgla w fotosyntezie. Szlak fotosyntezy C3 różni się chemicznie od szlaku C4, więc rośliny C3 mają wykrywalnie różne proporcje izotopów węgla niż rośliny C4. Rośliny C3 to głównie drzewa, krzewy, zioła i trawy umiarkowane . Rośliny C4 obejmują kukurydzę, niektóre proso, sorgo i ciepłe trawy. Zwierzęta, które jedzą te różne rośliny, wykazują różne proporcje. Na przykład żyrafy to przeglądarki, które zjadają głównie rośliny C3, a zebry to zwierzęta żywiące się głównie roślinami C4. Dlatego skamieliny osobników, które mają wyższy stosunek izotopów węgla (C4), są oznaczone jako "pasażerowie", a te, które mają niski stosunek (C3), są odpowiednio przeglądarkami lub zjadaczami pasących się lub zjadaczy przeglądarek. Parantropus wykazuje mieszaną sygnaturę węglową, podobnie jak mrówki, które żywią się głównie termitami (które należą do sygnatury C4). Tak więc Parantropus, podobnie jak jego krewni szympansy, mógł włączyć termity do swojej przeciętnej diety. Analiza narzędzi kostnych z miejsc jaskiń w RPA potwierdza tę tezę. Narzędzia są niezwykle podobne do eksperymentalnie wykonanych różdżek termitowych z wypolerowanymi, zaokrąglonymi końcówkami i równoległymi mikroskopijnymi prążkami z gleby. Izotopy węgla wskazują również, że Paranthropus zjadał tropikalne trawy i turzyce, drzewiaste owoce, krzewy i zioła. Różne sygnatury, które pojawiają się naprzemiennie w zębach, wskazują, że ich dieta była sezonowa i zmieniała się między różnymi roślinami w zależności od pory roku. Oczywiście, bezpośrednie dowody na dietę można znaleźć w zawartości żołądka danej osoby, jak również w skamieniałych odchodach (koprolitach) i wymiocinach. Nasiona w koprolitach homininów z Terra Amata (380 Kya) sugerują, że kemping był sezonowy, ponieważ poszczególne nasiona były dostępne tylko latem. Niestety zwierzęta zwykle nie ulegają fosylizacji z zachowanymi narządami. Gdyby zwierzęta kopalne zachowały żołądki, zawartość żołądków byłaby bezpośrednim dowodem na dietę, a przynajmniej na ostatnią kolację. Słynne ciała bagienne Wielkiej Brytanii i Danii często miały w żołądkach konserwowane pełne posiłki. Choroby związane z żywnością mogą również powiedzieć wiele o diecie danej osoby. Tak jest w przypadku KNM-ER 1808, częściowego szkieletu dorosłej samicy H. erectus z Koobi Fora w Kenii. Jej długie kości były pokryte tkaną kością, co jest oznaką choroby. Kość powinna być gładka, nie wyboista jak jej, a patologia jest zgodna ze współczesnymi przypadkami hiperwitaminozy A, choroby wywoływanej u współczesnych ludzi przez przedawkowanie witaminy A. Jednak we wczesnym plejstocenie choroba była prawdopodobnie spowodowana jedzeniem zbyt wielu wątrób mięsożernych, ponieważ wątroby mięsożerców są bogate w witaminę A w porównaniu z innymi dostępnymi na sawannie wschodnioafrykańskiej. Jeśli H. erectus był w stanie przedawkować produkt zwierzęcy około 1,5 miliona lat temu, jasne jest, że do tego czasu potrafił zdobyć mięso.
Tasiemce
Ludzki tasiemiec oferuje intrygujące spojrzenie na ewolucję mięsa. Zwierzęta domowe są od dawna obwiniane za dawanie ludziom tasiemców (Taenia). Kiedy więc przeprowadzono analizy genetyczne tasiemców pochodzących od różnych żywicieli zwierzęcych, naukowcy spodziewali się znaleźć pochodzenie ludzkiego tasiemca mniej więcej w czasie, gdy rolnictwo stało się powszechne około 10 000 lat temu. Wręcz przeciwnie, wyniki pokazały, że tasiemce specyficzne dla człowieka nie są potomkami tasiemców żyjących wśród współczesnych gospodarzy zwierząt domowych. Ludzkie tasiemce ewoluowały dalej, wraz z przejściem do zachowań związanych z jedzeniem mięsa, gdy ludzie dołączyli do gildii mięsożerców. Cykl życiowy tasiemca obejmuje dwóch różnych żywicieli. Najpierw składa jaja na ziemi, które są zjadane przez zjadacza (jak antylopa), który jest "żywicielem pośrednim" tasiemca. Jajo tworzy zarodek w mięśniu zwierzęcia. Kiedy mięsożerca lub "żywiciel ostateczny" zjada żywiciela pośredniego, połyka młodego tasiemca, który wyrasta na dorosłego tasiemca i pozostawia mięsożercę z kałem. Tasiemiec został zatem przetransportowany z powrotem na ziemię, gdzie będzie składał jaja i kontynuował cykl życiowy. Sekwencje genów tasiemca są najbardziej podobne u ludzi, hien i dużych kotów w Afryce, więc homininy prawdopodobnie wybrały tasiemce z jedzenia pośrednich żywicieli tasiemców specyficznych dla tych drapieżników prehistorycznej afrykańskiej sawanny. Zegary molekularne sekwencji wskazują na pochodzenie około 1,71-0,78 Mya, czyli w czasach H. erectus.
Początki Człowieków (52)
UŻYWANIE NARZĘDZI
Ludzie nie są jedynymi zwierzętami, które używają narzędzi. Wydry morskie, nagie kretoszczury i sowy to tylko niektóre z innych użytkowników narzędzi na naszej planecie. Matki delfinów uczą córki używania gąbek do ochrony pysków przed kąsającymi stworzeniami podczas żerowania na dnie morskim. U naczelnych zwyczajowe używanie narzędzi (takich jak używanie młotków, sond i skrobaków wykonanych ze skał lub roślin) ogranicza się głównie do orangutanów, szympansów , bonobo, goryli i kapucynek (małpy Nowego Świata). Goryle i szympansy (zwłaszcza z lasu Tai w Wybrzeżu Kości Słoniowej) rozłupują orzechy palmy olejowej, rozbijając je między dwiema skałami młotkiem i kowadłem. Zaobserwowano, że goryle używają patyków i gałęzi jako kul do przechodzenia przez wodę, a szympansy używają liści jako gąbek do zbierania wody pitnej. Istnieje jednak różnica między używaniem znalezionego przedmiotu jako narzędzia a modyfikacją przedmiotu do użycia jako narzędzia, co robi niewiele zwierząt, poza szympansami i ludźmi. Samice szympansów i ich córki przygotowują patyk, a następnie zanurzają go w kopcu termitów, aby wydobyć pożywne owady. Szczotkopodobną końcówkę sondy do połowu termitów wykonuje się poprzez przeciąganie łodygi rośliny przez zaciśnięte zęby trzonowe, prostowanie jej do punktu poprzez zwilżenie jej śliną, a następnie ostrożne manipulowanie włóknami ręcznie lub ustami. Kije i gałęzie służą również do zabijania znacznie większej zdobyczy. Zaobserwowano, że szympansy używają przygotowanych gałęzi jako włóczni do zabijania młodych krzewów. Z pewnością wczesne homininy używały materiałów roślinnych jako narzędzi, ale te artefakty nie zachowują się jak narzędzia kamienne. Ponieważ szympansy nie wytwarzają narzędzi kamiennych na wolności - to znaczy nie odcinają kawałków z zamiarem użycia nowo zmodyfikowanych narzędzi skalnych - artefakty kamiennych narzędzi przypisuje się rękodziełu naszych przodków homininów. Nawet kiedy kapucyny i szympansy uczą się robić kamienne narzędzia w niewoli, nie produkują nawet najprostszych narzędzi podobnych do Olduwańczyków. Siła poznawcza determinuje zdolność wytwarzania narzędzi kamiennych, podobnie jak anatomia nadgarstka i dłoni. Istnieje kilka kości dłoni i nadgarstków australopitów i wykazują one cechy pochodne, takie jak giętki nadgarstek, długi kciuk i szerokie opuszki palców, które są związane z ludzką manipulacją i zdolnością do wytwarzania kamiennych narzędzi. Co zaskakujące, dowody dotyczące powiększonego mięśnia pośladkowego wielkiego u ludzi również przyczyniają się do naszego zrozumienia użycia narzędzi homininowych. Mary Marzke i jej koledzy zwrócili uwagę, że Australopithecus liium z Afryki Południowej pokazuje, że mięsień pośladkowy wielki, który przyczepił się do niego w życiu, był duży i podobny do tego u ludzi. W porównaniu z szympansami układ ludzkiego pośladka maksymalnego zapewnia mechaniczną przewagę podczas używania narzędzia. Nasz gluteus maximus kontroluje rotację i ruchy tułowia, co utrzymuje nas w równowadze i stabilności, gdy używamy narzędzi, rzucamy przedmiotami, uderzamy pałkami, kopiemy i zasadniczo robimy wszystko, co wymaga energicznych ruchów ramion. Tak więc z rąk i pośladków wynika, że od 3 milionów lat australopity miały ciało do poważne i przyzwyczajone do używania narzędzi. Ale znowu, dwunożność i używanie narzędzi muszą być połączone, ponieważ mięsień pośladkowy wielki jest również ważny dla biegania. Ponieważ bieganie (w przeciwieństwie do chodzenia) jest serią skoków na jednej nodze, mięśnie pośladkowe są bardzo zaangażowane w tę aktywność. Narzędzia kamienne są znalezione w powiązaniu z dowodami na zachowanie homininów w jedzeniu mięsa. We wczesnym plejstocenie istnieją takie miejsca jak Olorgesailie, w Kenii, które pokazują, jak ważne stały się kamienne narzędzia. W pobliżu znajdują się setki ręcznych siekier i innych narzędzi kamiennych, które są związane z pozostałościami H. erectus. Wszechobecność ręcznych toporów i ich spójność kształtu w przestrzeni i czasie (ponad milion lat!) nasuwa pytanie o ich funkcję. Z pewnością służyły, jak sama nazwa wskazuje, jako podręczne siekiery do skórowania i uboju zwierząt. Dodatkowo płatki, które odpadły z nich podczas produkcji, byłyby również przydatne do cięcia narzędzi. Ale byłyby również przydatne do kopania bulw lub być może do kopania podziemnych źródeł wody. Jedna z hipotez wzywa ich aerodynamiczną naturę do postulowania, że zostały wystrzelone na zdobycz jako "zabójcze frisbee". Taki scenariusz łatwo sobie wyobrazić po wyglądzie Olorgesailie.
Początki Człowieków (51)
ZĘBY
Ponieważ szkliwo zębów jest wykonane z wytrzymałego materiału (hydroksyapatyt), zęby są dobrymi kandydatami do fosylizacji. W konsekwencji zęby są najczęstszą częścią szkieletów homininów, które są odkrywane i zbierane. Zęby można rozróżnić gatunkowo, ponieważ ich kształt i rozmiar są powiązane z wielkością ciała, dietą (do ścinania, ścinania, miażdżenia, szlifowania) i zachowaniami społecznymi (długie, ostre kły do współzawodnictwa partnerskiego). Różnice w liczbie, wielkości, kształcie, wzorze guzka i rozmieszczeniu zębów umożliwiają identyfikację gatunków przodków i spokrewnionych oraz śledzenie ewolucji diety. Jeśli uśmiechasz się do lustra, łatwo zauważyć, że w ustach człowieka są różne rodzaje zębów. Siekacze tnące z przodu są jak płaskie łopaty, kły przypominają małe kły lub dodatkowe siekacze, przedtrzonowce (które niektórzy dentyści nazywają "przedtrzonowcami"), a trzonowce są wyboistymi, wielofunkcyjnymi narzędziami do szlifowania. Ten stan posiadania zróżnicowanego zestawu narzędzi dentystycznych nazywa się heterodoncją. Skrajnie odwrotny stan, zwany homodoncją, obserwuje się w szczękach krokodyla, gdzie wszystkie zęby są ostrymi stożkami używanymi do chwytania i rozrywania. Wszystkie naczelne są heterodontyczne, ale można je odróżnić na podstawie określonych wzorów zębowych, które prezentują, oraz określonych kształtów i rozmiarów zębów. Ludzie mają uogólnione uzębienie wskazujące na wszystkożerne jak suidy (świnie i ich przodkowie). W rzeczywistości czasami trzeba dobrze wyszkolonego paleontologa, aby odróżnić skamieniałość suida od skamieniałości człekokształtnego człekokształtnego, jeśli została złamana lub zużyta w wyniku użytkowania w życiu. Podczas ewolucji homininów zęby kłów stawały się coraz mniejsze. Ogólnie rzecz biorąc, ludzie mają małe zęby jak na wielkość głowy i ciała oraz mają bardzo mały dymorfizm płciowy zębów w porównaniu z małpami człekokształtnymi i małpami. Wczesne homininy dostają coraz mniejsze kły, zanim pojawią się dowody na jedzenie mięsa (około 2 milionów lat temu), a ten trend wskazuje, że zmiany społeczne były prawdopodobnie głównym motorem redukcji psów, a nie zmiany dietetyczne. Ponieważ wielkość kła straciła przyczepność w rywalizacji partnerskiej, albo (1) rywalizacja o partnera zmniejszyła się, jak u gibonów, które tworzą monogamiczne wiązania par, albo (2) zapanowały inne formy rywalizacji o partnera, które miały niewiele wspólnego z bronią zębową. Biomechanika zębów i otaczającego nas narządu żucia jest zwykle skorelowana z rodzajem przetwarzanej żywności. Paranthropus miał duże, płaskie, grubo emaliowane zęby trzonowe i towarzyszący im zestaw silnych adaptacji żucia w kościach czaszki. Zęby najwyraźniej nie były przystosowane do ścinania trawy i wydają się być przystosowane do obsługi twardych pokarmów roślinnych, takich jak twarde nasiona i orzechy. Ludzie doświadczają mikroskopijnego urazu zębów z pożywienia, a te blizny spowodowane pożywieniem mogą być wykorzystane do wnioskowania o diecie homininów kopalnych. Wgłębienia i zadrapania na zębach spowodowane spożywaniem różnego rodzaju pokarmów mogą być powiązane z pewnymi rodzajami pokarmu w oparciu o rozmiar, intensywność, częstotliwość i lokalizację (np. siekacze kontra trzonowce). Pierwsze badania mikrozużycia przeprowadzono w latach 70. XX wieku za pomocą skaningowego mikroskopu elektronowego (SEM), ale trzydzieści lat później preferowanymi narzędziami są skaningowe mikroskopy laserowe. Zęby Paranthropusa wykazują mikrozużycie spowodowane twardym, ale nie ściernym pokarmem, który został określony jako orzechy lub nasiona z twardymi osłonkami, co potwierdza przydomek "Dziadek do orzechów", który Louis Leakey nadał czaszce OH 5 Paranthropus boisei.
Zęby mądrości
Trzecie zęby trzonowe nazywane są "zębami mądrości", ponieważ wyrzynają się na początku dorosłości między 18 a 22 rokiem życia. Wiele osób nie ma wystarczająco dużych szczęk, aby pomieścić trzecie zęby trzonowe i mogą zostać zaklinowane lub utknąć w szczęce, gdy nadejdzie czas, aby wybuchły. Osoby mające dostęp do opieki dentystycznej często usuwają trzecie zęby trzonowe, aby uniknąć bólu, infekcji i stłoczeń zębów, co prowadzi do wad zgryzu, większego bólu i trudności w żuciu. Od dawna zakładano, że takie problemy są wynikiem drobno przetworzonej miękkiej żywności, która zastąpiła diety wielu społeczeństw opartych na rolnictwie, począwszy od około 10 000 lat temu. Spożywanie miękkich pokarmów przez dłuższy czas w dzieciństwie nie stymuluje szczęk do wzrostu tak dużych, jak u tych, którzy jedzą twardą lub gruboziarnistą dietę, co powoduje wzrost kości i podparcie się sił. Jednak jeden szkielet skamieniałości z miejsca we Francji jest sprzeczny z teorią rolnictwa dotyczącą problemu zębów mądrości. "Dziewczyna Magdaleny" 15 Kya ze schroniska skalnego Cap Blanc wbiła się w zęby mądrości, ale przeszły one niezauważone (dlatego nazywano ją "dziewczyną"), dopóki potężne obrazowanie rentgenowskie nie pozwoliło naukowcom zobaczyć ich w ich kryptach w jej szczęka. W chwili śmierci okaz był w wieku od dwudziestu pięciu do trzydziestu pięciu lat i powinien być teraz nazywany "Kobietą Magdaleną". Ponieważ jest ona starsza od najwcześniejszych dowodów na rolnictwo, miękka przetworzona żywność niekoniecznie jest winna jej problemu. Możliwe jest jednak, że ludzie przed rolnictwem przetwarzali dziką żywność w sposób podobny do tego, w jaki później ludzie zaczęliby wkrótce przetwarzać udomowioną żywność. Dalsze dowody archeologiczne rzucą na to światło w przyszłości.
Początki Człowieków (50)
WIELKOŚĆ, KSZTAŁT I WYTRZYMAŁOŚĆ CIAŁA
Wiele ludzkich adaptacji postępuje według tych samych wzorców, co inne ssaki na całym świecie, zwłaszcza te, które są determinowane przez zasady termoregulacji. Istnieją ogólne relacje obejmujące cały ssak między powierzchnią ciała a objętością (SA/V), które są kontrolowane przez klimat. Reguła Bergmana mówi, że ssaki w chłodniejszym klimacie mają zwykle większe ciała niż zwierzęta w cieplejszych. Większy rozmiar ciała zmniejsza stosunek SA/V, a tym samym zmniejsza straty ciepła. Opierając się na podobnej potrzebie zachowania ciepła, Reguła Allena stwierdza, że ssaki w chłodniejszym klimacie mają zwykle krótsze wyrostki niż zwierzęta w cieplejszych. Długości odcinków dystalnych, dłoni, przedramion, stóp i goleni są krótsze w populacjach ludzi żyjących w chłodniejszych środowiskach na wyższych szerokościach geograficznych, takich jak Eskimosi z Arktyki, w porównaniu z populacjami żyjącymi w gorących regionach w pobliżu równika, podobnie jak ludy Nilotów w Sudanie, których ciała są zbudowane tak, aby rozpraszały ciepło. Wraz z pojawieniem się H. erectus we wczesnym plejstocenie, wzrostowi wielkości ciała homininów towarzyszy liniowość (tj. chudość) budowy ciała, aby utrzymać stosunek SA/V odpowiedni dla gorącego, suchego klimatu.

Dłuższe ręce i nogi oraz smukły tułów w stosunku do wielkości ciała zapewniają większy potencjał chłodzenia poprzez pocenie się. Chłopiec Nariokotome H. erectus był liniowy jak dzisiejsze populacje nilotów, a może nawet bardziej. Korzyści płynące z budowy liniowej giną w wilgotnych, zalesionych, zamkniętych środowiskach. Australopity o małych ciałach, podobnie jak populacje ludzi karłowatych, prawdopodobnie były w stanie zamieszkiwać wilgotne, zamknięte środowiska, a także otwarte, suche środowiska, ale H. erectus prawdopodobnie wolał otwarte, suche środowiska. Australopithecus garhi przedstawia mozaikę proporcji kończyn australopita i H. erectus z długimi rękami i długimi nogami, więc reprezentuje formę pośrednią między tymi dwiema grupami. Neandertalczycy wykazują przeciwieństwo budowy ciała H. erectus. Mieli ekstremalną adaptację do niskich temperatur, nawet w porównaniu z najbardziej przystosowanymi do zimna ludźmi. Dzięki stosunkowo krótkim rękom i nogom oraz grubym torsom o baryłkowatym torsie są klasycznymi przykładami Reguł Bergmana i Allena. Samce neandertalczyków miały średnio około 5 stóp 6 cali wzrostu i 180 funtów, a samice około 5 stóp i 1 cal wzrostu i 160 funtów. Chociaż ich wzrost był niższy, ich ciała były szersze, grubsze, mocniejsze i bardziej zwarte niż nasze. Chociaż ludzie są wyżsi i więksi niż wielu naszych krewnych naczelnych, w porównaniu z nimi jesteśmy słabi. Szympansy są cztery razy silniejsze od ludzi i jest to widoczne w ich kościach. Infrastruktura szkieletowa silnego ciała jest inna niż słabego. Kości, które zakotwiczają mocne i duże mięśnie, wykazują większe miejsca przyczepu mięśni, są grubsze i cięższe, mogą mieć pewne krzywizny, aby lepiej opierać się siłom, a także mają kościste podpory i rozpórki wewnątrz nich, które są widoczne w promieniowaniu rentgenowskim. Kości nieustannie się przebudowują w odpowiedzi na siły generowane podczas aktywności fizycznej (zasada znana jako "prawo Wolffa"), dlatego ćwiczenia są niezbędne do wzmocnienia kości, aby uniknąć szkodliwego poziomu utraty kości w późniejszym życiu. Osteoblasty to komórki progenitorowe, które tworzą kości, a osteoklasty są ich resorpcyjnymi odpowiednikami. Gruba zewnętrzna warstwa kości (kość korowa) reaguje na stres poprzez odkładanie większej ilości kości tam, gdzie jest poddawana siłom i resorpcję kości tam, gdzie nie jest obciążona. Na przykład kość ramienna w ramieniu dzierżącym rakiety elitarnych tenisistów, zwłaszcza tych, którzy zaczęli grać jako dzieci, jest znacznie grubsza niż druga ręka. Niewykorzystane kości będą ulegać resorpcji i atrofii zamiast przebudowywać nową kością jak zdrowa kość. Kości astronautów doznają poważnego urazu po spędzeniu dużej ilości czasu w zerowej grawitacji na pokładzie stacji kosmicznej, a naukowcy opracowują wygodne sposoby na przywiązanie astronautów do bieżni, aby dodać siły do ich kości podczas ćwiczeń. Wskaźniki siły kości można również śledzić w zapisie kopalnym homininów i jasne jest, że skamieniałe homininy były znacznie silniejsze i bardziej aktywne niż współcześni ludzie. Zwłaszcza w czasach H. erectus, kiedy rozmiary i proporcje ciała były podobne do naszych, widać wyraźnie, że kości były znacznie grubsze, a miejsca przyczepów mięśniowych również znacznie większe. Neandertalczycy są ekstremalnym przykładem wytrzymałości szkieletowej dzięki swoim grubym, ciężkim i czasami zakrzywionym długim kościom. Ale nawet zanim niektóre współczesne populacje ludzkie zaczęły prowadzić osiadły tryb życia z rolnictwem, ich ciała były stosunkowo słabsze niż ciała poprzednich homininów. Innowacje kulturowe prawdopodobnie zrekompensowały potrzebę H. sapiens aby utrzymać siłę fizyczną.
Początki Człowieków (49)
Zredukowane owłosienie na ciele
Ludzie mają drastycznie zredukowane owłosienie ciała, mimo że jest to ważne dla ochrony przed klimatem oraz ostrymi zębami i pazurami innych zwierząt. Futro na ciele może być również używane do sygnalizowania kolorem i wzorami, a także poprzez piloerekcję lub podnoszenie włosów, aby nadymać się i wyglądać na dużego i agresywnego. Ludzie nie tylko są funkcjonalnie "nadzy", ale też się pocimy. Mamy więcej gruczołów potowych niż jakikolwiek inny naczelny i jesteśmy również jednymi z najbardziej spoconych ssaków. Wynika z tego, że prawdopodobną przyczyną utraty włosów na ciele była potrzeba wzmożonego pocenia się. Nie ma dowodów kopalnych na ewolucję włosów na ciele człowieka, więc genetyka i adaptacje innych ludzkich cech kierują się hipotezami. Nina Jablonski przekonuje, że potrzeba termoregulacji podczas chodzenia z dużym, gorącym mózgiem w słonecznych, gorących okresach dnia (przypuszczalnie podczas żerowania, polowań i padlinożerstwa) doprowadziło do selekcji pod kątem utraty futra u wczesnych członków rodzaju Homo około 2 milionów lat temu. Utracie sierści towarzyszyła ewolucja większej liczby gruczołów potowych, które powodują obfite pocenie się (a tym samym chłodzenie ciała) oraz ewolucja ciemniejszej skóry chroniącej przed szkodliwym promieniowaniem ultrafioletowym (UVR). Połączenie utraty futra na ciele, zwiększonej tolerancji ciepła poprzez pocenie się i ciemniejszej skóry pozwoliło homininom podróżować dalej i spędzać dłuższe okresy czasu pod gorącym słońcem. Gen MCIR związany z pigmentacją skóry wskazuje na datę pojawienia się ciemnej skóry 1,7 miliona lat temu, co jest zgodne z hipotezą, że utrata futra na ciele i ciemna pigmentacja ewoluowały wspólnie około 2 milionów lat temu. Włosy są obfite na czubku głowy, gdzie słońce bezpośrednio pada na ciało, a także pod pachami i okolicami łonowymi, prawdopodobnie w celu ochrony przed otarciami i obiciami lub do przechowywania zapachów sygnalizujących płodność i tożsamość. Wszystkie włosy na ciele są właściwie tylko jednym z dwóch rodzajów: włosy końcowe występują na głowie, brwiach i rzęsach, ale włosy welusowe występują wszędzie indziej. Te typy włosów różnią się pod względem trwałości: wszystkie rosną w tempie około pół cala na miesiąc, ale mają różny czas wzrostu, zanim wypadną ze skóry. Genomika porównawcza pewnego dnia wyjaśni historię genetyczną stojącą za naszym unikalnym rozmieszczeniem włosów na ciele. Aby zbadać utratę futra ciała, możemy poszukać genów kandydujących w genomach bezwłosych myszy, nagich golców, bezwłosych kotów i psów (oczywiście z futrzastymi grupami zewnętrznymi dla porównania), aby sprawdzić, czy ludzie mają coś podobnego. Jeden gen kandydujący dla długich włosów jest znany z myszy (zwanych "myszami angora") z mutacją w FGF5, genie, który w swoim normalnym stanie zatrzymuje wzrost włosów. Być może ludzie mają również genetyczny mechanizm ignorowania genu zatrzymującego porost włosów. Istnieje również kilka genów keratyny, substancji tworzącej włosy. Wiele z tych genów jest identycznych u ludzi, szympansów i goryli, ale jeden jest inny u ludzi. Nie pełni żadnej widocznej funkcji u ludzi, ale koduje białka u innych naczelnych. Mutacja u ludzi miała miejsce około 250 Kya, co mogło być czasem, kiedy ludzkie włosy na głowie przybrały swoją obecną, stale rosnącą formę. Konieczne będą dalsze badania nad tymi genami, aby lepiej zrozumieć ten proces. Skoro owłosienie ciała zostało zredukowane, dlaczego owłosienie głowy zostało zwiększone? To może być ochrona mózgu przed bezpośrednim działaniem promieni słonecznych. Może być wybrany seksualnie ze względu na urodę. Może być wskaźnikiem kondycji zdrowotnej i społecznej. Ludzkie włosy wymagają dodatkowej pielęgnacji, a poświęcenie czasu na pielęgnację włosów lub kogoś innego może sygnalizować ich znaczenie lub status. Figurka Wenus z Willendorfu w Austrii miała wyszukaną fryzurę, pokazującą nam wczesne spojrzenie na znaczenie dobrych włosów przez 23 Kya.
Początki Człowieków (48)
DWUNOŻNOŚĆ
Ludzie są jedynymi ssakami, które zwykle chodzą i biegają na dwóch nogach. Jednym z pytań wartych milion dolarów w pochodzeniu człowieka jest to, dlaczego czworonożna małpa żyjąca w wieku około 6 milionów lat zaczęła chodzić na dwóch nogach? Cztery nogi są znacznie lepsze niż dwie pod względem siły, szybkości i równowagi. Poza tym, jakiego rodzaju presja selekcyjna była potrzebna, aby ewoluować adaptacje do dwunożności od czworonożności? Drugim pytaniem, które idzie w parze z pytaniem "dlaczego" jest pytanie "kiedy" - kiedy homininy stały się nawykowo dwunożne i straciły większość swoich nadrzewnych adaptacji? Przedstawiono kilka hipotez wyjaśniających pochodzenie dwunożności, które jest również zwykle uważany za klin, który dzieli LCA na dwie grupy: homininy i linię, która doprowadziła do powstania szympansów. Wiele z nich wyjaśnia utrzymywanie się selekcji w przypadku dwunożności po jego wyewoluowaniu, ale niewielu wyjaśnia samo jego pochodzenie lub impuls do jego ewolucji. Energetyka mogła odegrać pewną rolę. Dwunożność człowieka jest skuteczna; chodzenie w pozycji wyprostowanej zajmuje człowiekowi mniej energii niż szympansowi chodzenie na czworakach na tej samej odległości. Być może dzieje się tak dlatego, że szympansy mają anatomię, która pozwala im zarówno podróżować po ziemi, jak i wspinać się po drzewach w poszukiwaniu owoców, ale ludzie specjalizują się wyłącznie w ruchu naziemnym. Ale kiedy szympansy są szkolone do chodzenia na dwóch nogach, nie chodzą wydajnie. Czapią się, walczą o równowagę i nie mają naturalnie gładkiego chodu ani wymachu ramion. Przypuszczalnie ta trudność byłaby podobna do tej, której doświadczały pierwsze dwunożne. Zatem selekcja musiała być bardzo silna we wczesnych fazach dwunożności, aby przezwyciężyć trudność balansowania na dwóch nogach. Co mogło skłonić selekcję do pokonania początkowych przeszkód anatomicznych? Termoregulacja, a konkretnie potrzeba unikania przegrzania, mogła być przyczyną dwunożności ewolucji. Pomysł ten wskazuje, że lepiej chodzić wyprostowany w stresującym upale południowego słońca na suchej sawannie wschodnioafrykańskiej niż na czworakach, ponieważ wyprostowana postawa oznacza, że mniej skóry jest narażonej na bezpośrednie działanie promieni słonecznych. Duży ludzki mózg jest wrażliwy na stres cieplny, więc taka strategia mogła być korzystna. Dodatkowo istnieją inne adaptacje dedykowane do chłodzenia ciała, które wskazują na jego znaczenie, takie jak wzrost gruczołów potowych i utrata sierści ciała, które pomagają w wyparnym chłodzeniu skóry. Być może chodzenie dwunożne wyewoluowało z czujnej postawy wyprostowanej używanej do wypatrywania drapieżników z drzew lub podczas chodzenia między łatami drzew. Wysychanie Afryki Wschodniej doprowadziłoby do coraz bardziej nierównych lasów, a hipoteza ta jest powiązana z jedną z najstarszych, znaną jako "hipoteza sawanny", która twierdzi, że homininy musiały przenieść się na ziemię po prostu dlatego, że na wysychającej sawannie afrykańskiej było mniej drzew . Jednak teraz jest jasne, na podstawie paleośrodowiskowych rekonstrukcji miejsc, w których znaleziono skamieliny Sahelanthropus, Orrorin i Ardipitheucus, że najwcześniejsze homininy, które prawdopodobnie były nieco dwunożne, żyły w lasach. Stanie prosto uwolniło ręce od ich roli lokomotorycznej, a Darwin, Owen Lovejoy i inni podkreślali rolę manipulacji w swoich hipotezach dotyczących ewolucji dwunożnej. Szympansy i goryle mogą nosić przedmioty w jednej ręce, opierając je o biodro podczas chodzenia w technice "trójnogu", a także mogą przenosić przedmioty, wciskając je między żuchwę a klatkę piersiową. Ale dwunożne homininy używałyby obu rąk do robienia narzędzi i noszenia ich, a selekcja była zdecydowanie silna do manipulacji, ponieważ w czasach australopitów jasne jest, że ich dłonie rozwinęły zdolności precyzyjnego chwytania, które były lepsze niż u szympansów. Kciuk jest dłuższy, a jego staw jest bardziej ruchliwy jak u człowieka. Anatomiczne dowody na dwunożność wyprzedzają pierwsze narzędzia kamienne o co najmniej 2,5 miliona lat, ale użycie materiałów niekonserwujących mogło być równie znaczące (patrz sekcja "Używanie narzędzi" w tym rozdziale). Kolejną zaletą wolnych rąk byłoby noszenie żywności z dala od niebezpiecznych obszarów, takich jak zwłoki, w których czają się padlinożercy. Noszenie jedzenia potencjalnemu partnerowi w zamian za seks lub zaopatrzenie rodziny było również oferowane jako hipoteza dwunożności. Chodzenie na dwóch nogach może być po prostu przedłużeniem stania na dwóch nogach w celu karmienia z małych drzew lub postawy dwunożnej podczas karmienia na drzewach. Szympansy i goryle stoją dwunożnie, aby się pożywić, kiedy tego potrzebują, a ich niemowlęta są w rzeczywistości bardzo dwunożne i krzyczą wokół swoich matek, aby utrzymać równowagę, a z kolei ich matki czasami chodzą dwunożnie, aby je nosić. Bez względu na przyczynę i siłę napędową (może to być kombinacja wszystkich do pewnego stopnia w przestrzeni i czasie), uwolnienie rąk doprowadziło do głównych specjalizacji w ręce, nadgarstku i palcach w manipulowaniu przedmiotami i przenoszeniu wszystkich funkcji lokomotorycznych na nogi (poza używaniem ramiona do kołysania się na bok) doprowadziły do głównych specjalizacji w całym ciele do chodzenia i biegania tylko na dwóch nogach. Nie wiadomo dokładnie, w jaki sposób nastąpiło przejście od nadrzewności do dwunożności (stopniowo lub szybko), ale hipotezy dotyczące diety i używania narzędzi są najsilniejsze. Kiedy jednak dwunożność wyewoluowała, nie zniknęły żadne ślady nadrzewności. Między 6 Mya a 2 Mya homininy zachowały wiele cech związanych z nadrzewnością, takich jak długie ramiona, zakrzywione palce u rąk i nóg oraz krótkie nogi. Pytanie brzmi: czy echa nadrzewności pozostały aż do 2 milionów lat temu, ponieważ homininy nadal do pewnego stopnia były nadrzewne, czy też przystosowanie się do dwunożności zajęło tylko dużo czasu? Chociaż debata dotyczy plioceńskiego zapisu hominina, szkielet Australopithecus afarensis "Lucy" (AL 288-1) jest zwykle plakatem dla obu stron problemu, podobnie jak reszta gatunku, ponieważ zapis kopalny dla niego obecnie oferuje większość dostępnych dowodów na przejściową formę hominina. Naukowcy z jednego obozu, w którym znajduje się Owen Lovejoy, twierdzą, że Lucy była dwunożna, podobnie jak współcześni ludzie. Wskazują na szeroki, rozkloszowany kości biodrowej (tj. jej miednicę przypominającą miednicę), "kąt nośny" kości udowej (kąt kości udowej w kolanie) oraz położenie otworu wielkiego pod czaszką. Twierdzą, że odciski stóp Laetoli (które prawdopodobnie zostały wykonane przez A. afarensis) wyraźnie pokazują mniej rozbieżny duży palec u nogi, dobrze rozwinięty łuk (do przechowywania energii i pochłaniania wstrząsów, których nie ma u małp płaskonogich) oraz wyraźną depresję dla wpływu uderzenia piętą, po którym następuje oderwanie palca (wyjątkowo ludzki wzorzec chodu). Ale ci z przeciwnego obozu, jak Jack Stern i Randall Susman, twierdzą, że A. afarensis w dużym stopniu polegał na drzewach, aby przetrwać, i dlatego byli wykwalifikowanymi wspinaczami. Wskazują, że Lucy i jej gatunek mają podobieństwa do adaptacji wspinaczkowych i suspensyjnych u małp człekokształtnych, takich jak klatka piersiowa w kształcie lejka, długie zakrzywione paliczki na dłoniach i stopach, anatomia barku (łopatki) wspinacza, stosunkowo krótkie nogi i stosunkowo długie ramiona. Wraz z odkryciem większej liczby skamieniałości będzie można ustalić, czy australopity pozostawały przynajmniej w niepełnym wymiarze czasu mieszkańcami drzew w całym pliocenie, czy też nie, ale jest jasne, nawet w chwili obecnej skamieniałości, że ten w pełni nawykowy dwunożność i jego cały zestaw ludzkich adaptacji lokomotorycznych, dostarczonych przez 1,8 Mya wraz z H. erectus.
Hipoteza Wodnej Małpy
Co jakiś czas w mediach pojawia się hipoteza Aquatic Ape (AAH; zwana także Aquatic Ape Theory), ale w recenzowanym czasopiśmie naukowym opublikowano tylko jeden adres AAH. Pomysł, że ludzkość wyrosła z wodnej przeszłości, po raz pierwszy przedstawił Alister Hardy (1896-1985), członek Towarzystwa Królewskiego i oceanograf biologiczny. Następnie autorka Elaine Morgan podjęła ten pomysł i od tego czasu napisała kilka książek we wsparciu hipotezy Hardy′ego. Zwolennicy AAH wskazują, że skamieniałości homininów znajdują się w bliskiej odległości od wody, a często w pobliżu dużych zbiorników wodnych, takich jak jeziora Turkana, Tanganika i Wiktoria. Zakładają, że wodne lub przynajmniej półwodne pochodzenie może wyjaśniać główne różnice fizyczne między ludźmi a innymi naczelnymi, takie jak zdolność pływania, zdolność noworodków do pływania i unoszenia się na wodzie oraz względna bezwłosość naszych ciał, jak wiele innych ssaków wodnych. W rzeczywistości, uosobienie ludzkości, dwunożność i duży mózg, są również wyjaśnione przez AAH: aby przebrnąć przez wodę, homininy stały się lepszymi dwunożnymi, a następnie karmienie się rybami i skorupiakami dostarczało pożywienia do rozwoju dużego mózgu. Chociaż AAH często pojawia się w prasie, większość paleoantropologów uważa ją za taką historię z wielu powodów. Brodzenie i pływanie nie są charakterystyczne dla ludzkiej kondycji. Jeśli nadarzy się okazja, na wolności lub w niewoli, wiele innych naczelnych brodzi (nawet dwunożnie) i pływa. Samce goryla nizinnego srebrnogrzbietego pluskają się gwałtownie rękami na bagnach w symbolicznych pokazach, a inne wygodnie stoją i siadają, by pożywić się na bagiennych trawach. Wiadomo, że japońskie makaki odpoczywają w naturalnych gorących źródłach. Najwyraźniej naczelne inne niż ludzie są zdolne do rozwijania hydrofilnych zachowań, które nie skutkują dwunożnością, bezwłosością, rachunkiem różniczkowym czy poezją. Jednak ta linia dowodowa może być również wykorzystana przez zwolenników AAH, ponieważ jeśli powinowactwo do wody jest wspólne dla wielu wyższych naczelnych, prawdopodobnie było ono obecne we wczesnych homininach i mogło być siłą napędową ewolucji człowieka. Z pewnością w pobliżu wody znajduje się wiele skamieniałości homininów. Jednak skamieniałości są najczęściej zachowane w pobliżu wody, ponieważ są szybko zakopywane w osadach, a w większości przypadków woda jest transporterem tych osadów. Miejsca skamieniałości bez pochówku wodnego lub przynajmniej częściowej ingerencji wody są rzadkie, chyba że zostaną znalezione w ekstremalnych warunkach pustynnych lub arktycznych lub w przypadku natychmiastowego pochówku przez lawę wulkaniczną lub popiół. Paleontolodzy doskonale zdają sobie sprawę z błędu zachowania i oczekują, że skamieliny można znaleźć w pobliżu starożytnych rzek, strumieni, delt i jezior ze względu na korzyści, jakie zapewniają. Chociaż AAH nie jest silną ani mainstreamową hipotezą naukową, nie powinniśmy przeoczyć możliwości, że starożytne homininy zanurzały się, brodziły i tarzały, aby zachować spokój, tak jak my i wiele innych ssaków.
Początki Człowieków (47)
PSYCHOLOGIA EWOLUCYJNA
Czy kiedykolwiek czytałeś kolumnę z poradami dietetycznymi, która sugeruje, że twoja niezdolność do opierania się smażonemu w głębokim tłuszczu fast foodowi była w rzeczywistości adaptacyjna w twojej ewolucyjnej przeszłości? Jest to przykład popularnego zastosowania psychologii ewolucyjnej (EP), która jest połączeniem biologii ewolucyjnej i psychologii poznawczej. Dwóch najwybitniejszych psychologów ewolucyjnych, John Tooby i Leda Cosmides, postrzega ludzki umysł jako szwajcarski scyzoryk z setkami, a nawet tysiącami "modułów", które wykonują określone zadania. Moduły te, podobnie jak inne części ciała, ewoluowały na różnych etapach ewolucji homininów między LCA i przed nadejściem rolnictwa i cywilizacji około 10 Kya, kiedy wszyscy ludzie żyli w społeczności zbieraczy na małą skalę, takie jak współcześni łowcy-zbieracze. Przykłady modułów umysłu obejmują te dotyczące preferencji żywieniowych (takich jak tłuszcze i cukry), kojarzenia się, teorii umysłu, unikania drapieżników, tworzenia sojuszy i języka - z których wszystkie ewoluowały, aby rozwiązać problemy adaptacyjne, które były kluczowe dla przekazywania genów. Na przykład teoria modułu umysłu pozwala ludziom zrozumieć stany psychiczne innych. Dzieci przed osiągnięciem czterech i pół roku życia nie potrafią zrozumieć, że inni mogą wyznawać inne przekonania niż one i w rezultacie nie mogą kłamać przekonująco. Ta wyjątkowo ludzka, ale nie wyuczona cecha odgrywa kluczową rolę w empatii, oszukiwaniu (siebie i innych) oraz manipulacji w sytuacjach politycznych/społecznych. Zasady EP są często stosowane do naszych nawyków żywieniowych i picia oraz zaburzeń. Według EP, ludzie wyewoluowali moduły preferencji żywieniowych. Moduły te ewoluowały, aby przetrwać na bezlitosnej afrykańskiej sawannie i rządzą pragnieniem pewnych pokarmów i niechęcią do szkodliwych. Tłuszcz pozyskiwany z padlinożernych lub polujących zwierząt oraz cukier pozyskiwany z dojrzałych owoców dostarczają składników odżywczych, których na sawannie byłoby stosunkowo mało. Zarówno tłuszcz, jak i cukier są potrzebne do wzrostu, utrzymania i metabolizmu (produkcja energii). Zwłoki i drzewa owocujące są etapami rywalizacji międzygatunkowej i z pewnością byłyby niebezpieczne strefy interakcji między drapieżnikami a homininami lub między innymi gatunkami naczelnych a homininami. Aby utrzymać zachowanie, korzyści muszą przewyższać koszty. Tak więc według EP, moduły preferencji tłuszczu i cukru ewoluowały, aby sprawić, że jednostki pożądają pokarmów wystarczająco silnie, aby zachęcić ich do podejmowania ryzyka. Nie zaobserwowano, aby współcześni ludzie tracili tłuszcz i cukier, mimo że wielu ludzi żyje w środowisku, w którym tłuszcz i cukier są łatwo dostępne. Nasza ewolucyjna historia napotkała współczesne niedopasowanie środowiska. Teraz grozi nam przejadanie się tłuszczem i cukrem do tego stopnia, że otyłość i choroby serca są zabójcami numer jeden w krajach uprzemysłowionych. Napady objadania się, które są uważane za zaburzenia odżywiania, które prowadzą do otyłości i chorób serca, mogły być adaptacyjne w przeszłości, gdyby ludzie, podobnie jak inne duże drapieżniki, jedli duże posiłki w odstępach kilku dni. Jeden z argumentów w sprawie EP stawia tych, którzy widzą wszystko jako adaptacje, przeciwko tym, którzy widzą wiele cech jako zwykłe produkty uboczne ograniczonej liczby adaptacji. Na przykład, niektórzy badacze uważają, że uniwersalna, międzykulturowa wiara w nadprzyrodzone wyjaśnienia wydarzeń lub "siły wyższej" wynika z modułu, który jest na stałe wpisany we wszystkie nasze mózgi, ale inni uważają to za spin. - od innych funkcji związanych z wyższym poznaniem i świadomością.
Początki Człowieków (46)
Interpretacja dowodów
WIELKI MÓZG I INTELIGENCJA
Wielki ludzki mózg ewoluował stosunkowo późno w ewolucji homininów, gdy na scenie pojawił się Homo erectus. Ponieważ jednak ludzki mózg jest postrzegany jako mistrz ludzkiej ewolucji, rozważymy go najpierw. Niepohamowana ciekawość otaczająca ewolucję ludzkiego mózgu jest podtrzymywana przez samą materię, która jest tak zagadkowa. Jednak wewnątrz ludzkiej czaszki nie ma nic wyjątkowego. Jedynie względne rozmiary obszarów anatomicznych w mózgu oraz liczba neuronów i charakter ich sieci są unikalne dla ludzi. Naczelne na ogół mają duże mózgi w porównaniu z większością ssaków. W rzeczywistości neandertalczycy mieli nawet większe mózgi niż my, więc to okablowanie naszych mózgów (liczba i natura sieci neuronowych), a nie rozmiar naszych mózgów, odróżnia ludzi. W ludzkim mózgu istnieje specyficzny rodzaj neuronu "wrzeciono", które jak wiadomo, że istnieje tylko w mózgach małp człekokształtnych i waleni (rzędu ssaków, który obejmuje delfiny i wieloryby). Jak większość dna oceanicznego, w mózgu znajduje się wiele niezbadanych obszarów. Nadal nie wiadomo dokładnie, co robią neurony wrzeciona i jak (lub czy) korelują z wyższymi zdolnościami poznawczymi. Jednak prawdopodobnie nie jest przypadkiem, że neurony wrzecionowate są wspólne dla grup, które dzielą złożone wzorce społeczne, złożone umiejętności komunikacyjne, tworzenie koalicji, współpracę, transmisję kulturową i używanie narzędzi. Rozmiar mózgu jest skorelowany z inteligencją na poziomie gatunku. Ludzie mają największe mózgi jak na swój rozmiar ciała, a walenie i małpy człekokształtne są tuż za nimi, a te gatunki są uważane za najbardziej inteligentne zwierzęta. Wyższe poziomy inteligencji ssaków charakteryzują się elastycznym rozwiązywaniem problemów w złożonych scenariuszach, włączaniem nowatorskich rozwiązań do istniejących zachowań i ciekawością. Inteligencję ludzką wyróżnia na przykład skłonność do myślenia abstrakcyjnego i symbolicznego. Powiększonemu płatowi czołowemu człowieka w stosunku do małp człekokształtnych przypisuje się zaawansowane emocje, świadomość i pamięć. Udary do części płata czołowego mogą wpływać na procesy podejmowania decyzji moralnych. W szczególności obszar przedczołowy jest nieznacznie powiększony u małp człekokształtnych i ludzi i zawiera ośrodki przewidywania i planowania. Wiele z tego, co wiadomo o inteligencji małp, pochodzi z laboratorium, w którym zwierzęta są wprowadzane w sytuacje, których nie doświadczają na wolności. Szympansy spontanicznie rysują i malują w niewoli. Na Uniwersytecie w Kioto w Japonii szympans o imieniu "Ai" nauczył się ponad 100 symboli wizualnych, w tym leksygramów, japońskich znaków kanji, liter alfabetu, cyfr arabskich, a także nauczył się rozumieć ludzką mowę i gesty. Wiele innych szympansów i goryli w niewoli nauczyło się rozumieć ogromne słownictwo angielskich słów w kontekście złożonych struktur zdaniowych. Na wolności małpy wytwarzają i używają narzędzi. Wiadomo nawet, że szympansy oszczędzają narzędzie do późniejszego użycia, co wymaga ostrożności. W obrębie gatunku rozmiary mózgu są różne, a korelacja między rozmiarem mózgu a inteligencją nie utrzymuje się. Osoba z mózgiem na dolnym końcu spektrum zmienności niekoniecznie jest mniej inteligentna niż osoba z największym mózgiem na świecie. Problematyczne jest również mierzenie inteligencji. Na przykład testy ilorazu inteligencji (IQ) stosowane u ludzi mogą mierzyć tylko określone aspekty wyższych funkcji mózgu, a nie całe zjawisko. Inteligencja fizyczna, podobnie jak potężna i złożona koordynacja neuronalna, której wymaga rzucenie piłki baseballowej z prędkością 15 km/h w rękawicę łapacza, jest całkowicie ignorowana przez standardowe testy inteligencji. Bez możliwości łatwego pomiaru poziomu inteligencji w celu porównania między gatunkami, mierzalny rozmiar mózgu jest używany jako wskaźnik poziomu inteligencji. Zamiast używać podstawowego rozmiaru mózgu, porównuje się rozmiar kory nowej, która jest obszarem rozumowania przestrzennego i percepcji sensorycznej, i składa się z zewnętrznej powierzchni mózgu. W szczególności stosunek wielkości kory nowej do wielkości reszty mózgu koreluje z rozwiązywaniem problemów i elastycznością lub "inteligencją" u naczelnych. Robin Dunbar i współpracownicy wykazali, że wielkość kory nowej jest związana z wielkością grupy społecznej u naczelnych (przypuszczalnie dlatego, że złożona socjalizacja wymaga wyjątkowej inteligencji). Zapis kopalny pokazuje, że mózg hominina wyraźnie powiększa się z czasem, podobnie jak rozmiar ciała. Ponieważ wielkość mózgu i wielkość ciała są skorelowane między gatunkami ssaków, same wartości wielkości mózgu nic nie wskazują bez uwzględnienia wielkości ciała. Współczynnik encefalizacji (EQ) gatunku jest stosunkiem wielkości jego mózgu do oczekiwanej wielkości mózgu w oparciu o znaną korelację między wielkością mózgu a wielkością ciała u ssaków. EQ oferuje sposób porównywania rozmiarów mózgu między gatunkami o bardzo różnych rozmiarach (takich jak wieloryby i ludzie) oraz śledzenia wzrostu rozmiaru mózgu u homininów. Znaczący wzrost wielkości mózgu w stosunku do wielkości ciała wystąpił u H. erectus i trwał do pojawienia się współczesnych ludzi. Kilka różnych czynników mogło przyczynić się do wyboru większego mózgu w H. erectus i poza nim. Ale najpierw powinniśmy rozważyć ewolucję inteligencji naczelnych. Główne hipotezy dotyczące ewolucji inteligencji naczelnych opierają się na zdobywaniu pożywienia, ekstrakcji pożywienia i uspołecznianiu. Jeśli źródło pożywienia jest sezonowe, jak owoce, naczelny musi w myślach mapować miejsca, planować trasy oraz przewidywać dostępność i jakość zasobów. Takie potrzeby poznawcze mogą również wymagać cech takich jak widzenie ostre, stereoskopowe i barwne. Mangabey o szarych policzkach (gatunek małpy afrykańskiej) wyewoluowały w zdolność wykorzystywania pogody do przewidywania, kiedy figi dojrzeją. Jeśli pogoda była ciepła, wrócą na drzewo, na którym, jak wiedzą, dojrzewają figi. Pozyskiwanie pokarmów, takich jak twarde orzechy, korzenie, larwy owadów, strąki nasion i łodygi, wymaga wyższych funkcji poznawczych, a także precyzyjnych zdolności motorycznych. Życie w grupie, jak większość gatunków naczelnych, obejmuje rywalizację, konflikty, przynależność, współpracę, dobór krewniaczy i wzajemność oraz śledzenie zachowań innych. Inteligencja naczelnych może ewoluować i ewoluowała dzięki którymkolwiek z tych modeli lub ich kombinacji. Z pewnością te problemy występowały we wczesnych homininach i wszystkie z nich mogły wyjaśniać zwiększenie rozmiaru mózgu, ale ponieważ ludzkie mózgi są większe i zdolne do wyższych funkcji poznawczych niż inne naczelne, coś innego musiało mieć znaczenie. Tkanka mózgowa jest bardzo kosztowna w uprawie i utrzymaniu. Stworzenie tak dużych mózgów wymagałoby silnej selekcji. Z pewnością pomogłaby wzbogacona dieta bogata w składniki odżywcze, szczególnie bogata w niezbędne nienasycone kwasy tłuszczowe dostarczane przez białko zwierzęce. Jest prawdopodobne, że włączenie mięsa do diety - co zbiegło się ze wzrostem rozmiarów ciała i mózgu w czasach H.erectus - przyczyniło się do ewolucji rozmiaru mózgu. Nie wiadomo konkretnie, dlaczego selekcja sprzyjała wzrostowi wielkości mózgu, chociaż łatwo jest wymyślić powody związane z kulturą, dietą, socjalizacją, rozmnażaniem itp. Pewne role odgrywały prawdopodobnie geny mikrokefalina i ASPM. Mikrocefalina jest krytycznym regulatorem rozwoju mózgu, a mutacje w genie powodują zaburzenie zlokalizowane w mózgu - małogłowie, który jest poważnym zmniejszeniem rozmiaru mózgu, nawet o 70 procent. Funkcje mózgu są często zlokalizowane w lewej lub prawej półkuli, co nazywa się lateralizacją. Jedną z takich zlateralizowanych funkcji jest język, którego ośrodki znajdują się na lewej półkuli. W kilku książkach, począwszy od 1983 roku i trwających do dziś, William Calvin zaproponował, że przeważająca tendencja do bycia praworęcznymi (kontrolowanymi przez lewą półkulę), a także do bycia wysoce zręcznymi, jest skorelowana z gromadzeniem się neuronów w lewej. półkula podczas rozwoju języka w pętli sprzężenia zwrotnego, w której język i ręczność były coraz bardziej wyspecjalizowane w selekcji złożonych sieci neuronowych. Hipoteza Calvina rzuca światło na znaczenie ewolucji inteligencji fizycznej (np. zręczności i posługiwania się rękami), która często jest przyćmiona przez ewolucję inteligencji umysłowej (np. języka).
Początki Człowieków (45)
STAROŻYTNE DNA I GENOM NEANDERTALSKI
Gdy warunki przechowywania są odpowiednie, możliwe jest wyodrębnienie DNA ze starożytnych, a nawet skamieniałych szczątków zwierzęcych i roślinnych. Zimne , suche środowiska suche lub mokre beztlenowe są zazwyczaj najlepsze do zachowania starożytnego DNA lub aDNA. Ponieważ przeżycie starych cząsteczek DNA jest rzadkie, a te, które przetrwają, faktycznie ulegają z czasem degradacji, istnieje ogólna zasada, że próbki starsze niż 100 000 lat nie są żywotnymi kandydatami do ekstrakcji aDNA. (Potencjalnym wyjątkiem mogą być 400 000-letnie rośliny zamrożone w syberyjskim lodzie, które mogły zachować aDNA.) Zdegradowane sekwencje aDNA można porównać z blisko spokrewnionymi współczesnymi gatunkami, aby skorygować błędy sekwencjonowania, które występują, gdy nukleotydy ulegają degradacji, aby "wyglądały" jak inne nukleotydy . Klonowanie organizmu z aDNA w celu przywrócenia go do życia byłoby możliwe tylko wtedy, gdy zachowane jest całe DNA, a szanse na to są bardzo małe. Jak dotąd aDNA zostało wyekstrahowane z co najmniej sześciu neandertalczyków (z Niemiec, Chorwacji, Gruzji, Belgii, Francji i Hiszpanii) i co najmniej pięciu współczesnych skamielin ludzkich (z Czech, Francji i Włoch). MtDNA jest przydatne do badań aDNA ponieważ w komórce znajdują się tysiące mitochondriów zawierających mtDNA, a nie tylko jedno jądro z DNA na komórkę. Po degradacji mtDNA ma znacznie większą szansę na zachowanie niż jądrowe DNA. Małe fragmenty kości lub zębów muszą zostać wywiercone i zmielone w celu analizy, ale na szczęście technologia odlewania jest w takim punkcie, w którym można wykonać wysokiej jakości repliki okazów przed ekstrakcją aDNA. Jest to destrukcyjny proces, dlatego paleoantropolodzy nieustannie rozważają zalety zachowania skamielin i ich morfologii z jednej strony, a nie wydobywania cennej informacji genetycznej, którą posiadają. Na podstawie starożytnych analiz mtDNA wiemy, że neandertalczycy trzy do czterech razy bardziej różnią się od współczesnych ludzi niż współcześni ludzie różnią się od siebie. Ponadto zmienność genetyczna między neandertalczykami a współczesnymi ludźmi jest znacznie większa niż w przypadku samych współczesnych ludzi. Zróżnicowanie neandertalskiego aDNA ze skamielin z odległych stanowisk jest podobne do zmienności wśród współczesnych populacji ludzkich. Neandertalczycy znajdują się między szympansami a ludźmi i poza żywym i kopalnym zasięgiem współczesnego człowieka. Opierając się na częstotliwościach zegara molekularnego mitochondrialnego aDNA, linia neandertalska oddzieliła się o około 500 Kya od linii, która doprowadziła do powstania współczesnych ludzi. Neandertalczycy, zgodnie z ich genami, różnią się od współczesnych ludzi i nie są bardziej spokrewnieni ze współczesnymi Europejczykami niż jakikolwiek inny współczesny człowiek (co jest uderzeniem w hipotezę wieloregionalnego pochodzenia człowieka, patrz rozdział 6). Dlatego jest mało prawdopodobne, abyśmy pochodzili od neandertalczyków i mało prawdopodobne, że dzieliliśmy z nimi geny (tj. krzyżowaliśmy się z nimi). Powinniśmy chyba uważać neandertalczyków za naszych dalekich kuzynów. Metody analizy aDNA są wystarczająco zaawansowane, aby umożliwić naukowcom, takim jak Svante Pääbo, ekstrakcję DNA jądrowego od neandertalczyków. Mając wystarczającą ilość aDNA z wystarczającej liczby próbek, ostatecznie zrekonstruowane zostaną duże fragmenty genomu neandertalczyka. Jak dotąd samiec 38 Kya znaleziony w jaskini Vindija w Chorwacji wytworzył sekwencję około miliona par zasad jądrowego DNA, co stanowi około 0,03 procent genomu. Za pomocą jądrowego DNA można analizować funkcjonalne geny i chromosomy płci. Na przykład chromosom Y neandertalczyków znacznie różni się od chromosomów ludzi i szympansów w porównaniu z innymi chromosomami, co sugeruje, że przynajmniej w ostatnich czasach neandertalczyków doszło do niewielkiego krzyżowania. Szacuje się, że neandertalczycy dzielą 99,5% swojego genomu z ludźmi, ale ludzie różnią się od siebie tylko o 0,1%. Tak więc, ponieważ zmienność się nie nakłada, wspierane jest oddzielne oznaczenie gatunkowe neandertalczyków i ludzi. Podobnie jak badania mtDNA, genom jądrowy neandertalczyków nie posiada żadnych pochodnych alleli, które są specyficzne dla populacji europejskich, co oznacza, że neandertalczycy nie przyczynili się do europejskiej ewolucji człowieka. Wkrótce będzie można badać funkcjonalne i chore geny wymarłych gatunków. Możliwe będzie również sprawdzenie, czy neandertalczycy przyczynili się do powstania ludzkiego genomu, jak niektórzy naukowcy wysunęli hipotezę, że tak jest w przypadku genu mikrocefaliny. Najbardziej obiecującym aspektem projektu genomu neandertalskiego jest możliwość poszukiwania specjalnych genów w ludzkim genomie dla języka i sztuki - cech i zachowań, które są unikalne dla ludzi i nie zawsze uważa się, że istniały u neandertalczyków. Kandydatami na te geny byłyby te wspólne dla szympansów i neandertalczyków, z wyłączeniem ludzi. Co więcej, geny wspólne dla neandertalczyków i współczesnych ludzi, ale nie szympansów, mogą wskazywać na zmiany genetyczne, które wystąpiły wcześniej w ewolucji homininów.
Metody na starożytne DNA
Zanim aDNA będzie mogło zostać zsekwencjonowane, musi zostać sklonowane i zamplifikowane przy użyciu techniki zwanej PCR (reakcja łańcuchowa polimerazy), która jest tą samą techniką, którą stosuje się w badaniu ojcostwa. Tysiące kopii aDNA są potrzebne do wykonania technik laboratoryjnych stosowanych w sekwencjonowaniu, a PCR dostarcza te kopie. Wszystkie analizy aDNA muszą odbywać się w fizycznie odizolowanym miejscu pracy, aby uniknąć skażenia ludzi, co może skutkować amplifikacją DNA, które nie jest badanym aDNA. W celu wykrycia rzadkiego aDNA należy przeprowadzić wielokrotne ekstrakcje ze skamieniałości i wiele procedur PCR. Czynność tak prosta, jak obchodzenie się ze skamieniałością bez rękawiczek, może zanieczyścić okaz, dlatego analizy DNA skamieniałości, które przez wiele lat znajdowały się w zbiorach muzealnych, są traktowane ze szczególną starannością. Ze względu na wszystkie szanse na skażenie człowieka i ponieważ może być bardzo trudno określić, czy aDNA lub DNA genetyka jest amplifikowane i sekwencjonowane, konieczne są dodatkowe środki ostrożności. Na przykład wyniki powinny być powtarzalne z tych samych, a różne ekstrakcje aDNA okazu kopalnego i oddzielne próbki okazu powinny być ekstrahowane i sekwencjonowane w niezależnych laboratoriach
Początki Człowieków (44)
ADAPTACJA CZŁOWIEKA
Chociaż geny zaangażowane w ekspresję złożonych cech szkieletowych i zębowych, które śledzimy w zapisie kopalnym homininów, wciąż są identyfikowane, przykłady adaptacji sterowanej selekcją u żywych ludzi ilustrują niedawną i ciągłą ewolucję naszego gatunku oraz stanowią przykład szybkości którego ewolucja może przekształcić populacje ludzkie. Utrzymanie tak szkodliwego allelu dla anemii sierpowatej pokazuje, w jaki sposób ludzie mogą przystosować się do własnej biologii do walki z chorobami. Niedawna ewolucja tolerancji laktozy w dorosłość w różnych populacjach pokazuje, jak szybko mogą ewoluować adaptacje dietetyczne, a także jak łatwo zachodzi ewolucja zbieżna. Wybór różnych ilości pigmentacji skóry u ludzi jest uderzającym przykładem tego, jak ludzie przystosowują się do określonych środowisk.
Cecha sierpowata
Gen z dwoma lub więcej allelami, każdy o znacznej częstości, jest polimorfizmem genetycznym. W niektórych populacjach afrykańskich gen odpowiedzialny za morfologię czerwonych krwinek jest jednym z takich przypadków. Jeden allel (który nazwiemy "A") koduje ekspresję prawidłowych krwinek czerwonych, ale allel sierpa (który nazwiemy "S") koduje krwinki czerwone w kształcie sierpa. Allel S jest spowodowany mutacją punktową zmieniającą tylko jeden aminokwas. Oba allele współdominują, więc żaden z nich nie maskuje ekspresji drugiego. Osoby z dwoma allelami sierpowatymi, które są homozygotyczne (SS), cierpią na anemię sierpowatą, która jest śmiertelną chorobą, w której sierpowate krwinki czerwone są zasadniczo upośledzone i nie mogą prawidłowo wykonywać swoich obowiązków związanych z transportem tlenu. Ze względu na negatywne skutki allelu S można by się spodziewać, że występuje on z niewielką częstotliwością, ale może osiągnąć częstotliwość 20 procent lub wyższą w populacjach w tropikalnych regionach Afryki, Azji Południowo-Wschodniej i Indonezji, gdzie dominuje malaria. Populacje żyjące w regionach zaatakowanych przez malarię wyewoluowały w naturalny sposób odporności na malarię przy użyciu allelu S. Osoby niebędące nosicielami (AA) mają normalne czerwone krwinki, ale częściej umierają na malarię niż nosiciele alleli sierpowatych (AS), którzy są odporni na malarię. Pasożyt malarii (różne gatunki Plasmodium), który żyje w czerwonych krwinkach, w rzeczywistości powoduje pękanie czerwonych krwinek w kształcie sierpa, więc pasożyt nie może jeść, przetrwać i rozmnażać się. Nosiciele alleli sierpowatych mają kilka komórek w kształcie sierpa, ale nie na tyle, aby spowodować poważne problemy zdrowotne lub śmierć. Jednak osoby z homozygotycznym genotypem (SS) najprawdopodobniej umrą z powodu anemii sierpowatej. Afrykanie noszący allel sierpa, którzy zostali zabrani do Ameryki w handlu niewolnikami, nie mieli już dla niego adaptacyjnego zastosowania w kraju bez malarii, więc udana adaptacja w Afryce jest teraz nieprzystosowaniem w Ameryce. Historia komórek sierpowatych to potężna lekcja, że geny mogą być korzystne lub szkodliwe w zależności od środowiska.
Tolerancja na laktozę
Zdolność do trawienia laktozy w wieku dorosłym jest stosunkowo nową adaptacją u ludzi, która pojawiła się wraz z nadejściem rolnictwa około 10 000 lat temu. Laktoza jest składnikiem mleka i produktów mlecznych. Laktaza jest enzymem rozkładającym laktozę podczas trawienia, dzięki czemu może zostać wchłonięta przez jelita. Normalni lub "typu dzikiego" ludzie mają stan przodków, który polega na zaprzestaniu produkcji laktazy w wieku około dwunastu lat. Są złymi wchłaniaczmi laktozy lub nie tolerują laktozy, podobnie jak inne dorosłe ssaki. Jednak niektóre grupy europejskie i afrykańskie, które mają długą historię pasterstwa i udomowienia zwierząt, nadal produkują laktazę w wieku dorosłym i zachowują zdolność trawienia laktozy. Wcześni ludzie byli wyłącznie myśliwymi i zbieraczami aż do około 10 Kya, kiedy niektóre populacje zaadaptowały nowe sposoby pozyskiwania pożywienia: używając udomowienia, aby utrzymać źródło pożywienia blisko. Presja selekcji była silna na tych ludziach, którzy byli w stanie zmaksymalizować te zasoby zwierzęce poprzez trawienie swoich produktów mlecznych przez całe życie. Zdolność do trawienia laktozy w wieku dorosłym musiała być ogromną zaletą żywieniową w tych populacjach, ponieważ selekcja silnie ją faworyzowała. Na przykład prawie wszyscy dorośli Holendrzy i Szwedzi tolerują laktozę. Analizy genetyczne przeprowadzone przez Sarah Tishkoff i innych wykazały, że grupy afrykańskie i europejskie zbiegały się niezależnie w tej samej adaptacji. Każda grupa wykorzystuje inny wariant genu regulatorowego (tj. nieco inny wariant allelu zwanego "SNP" dla polimorfizmu pojedynczego nukleotydu) do kontrolowania genu produkcji laktazy, który nazywa się LCT. Jak dotąd istnieją cztery różne mutacje, które utrzymują włączony gen laktazy. Każdy allel występuje znacznie częściej w populacjach, które mają długą historię uzależnienia od domowych zwierząt mlecznych: (1) Holendrzy i Szwedzi spokrewnieni ze starożytnymi hodowcami bydła "Funnel Beaker" z północno-środkowej Europy, (2) Nilo -saharyjskojęzyczne grupy w Kenii i Tanzanii, (3) ludność Beja z północno-wschodniego Sudanu i (4) afro-azjatyckie grupy mieszkające w północnej Kenii. Ewolucyjna opowieść o tolerancji laktozy jest potężna, ponieważ podkreśla wpływ kultury na ewolucję biologiczną. Co ważniejsze, trawienie laktozy jest doskonałym przykładem tego, jak szybko biologia człowieka może się przystosować, aby lepiej przetrwać.
Kolor skóry
Ze względu na szerokie spektrum zmienności, kolor skóry jest prawdopodobnie najczęstszą i najbardziej rzucającą się w oczy cechą, której ludzie używają do identyfikacji głównych różnic fenotypowych u innych. Jednak Nina Jablonski i jej koledzy wykazali, że za zróżnicowaniem koloru skóry człowieka kryje się silna funkcja adaptacyjna. Nasi najbliżsi krewni nie mają takiej odmiany koloru skóry jak nasza. Szympansy mają jasną skórę pod czarnym futrem. Młode szympansy mają jasne twarze, ale wraz z wiekiem i latami przebywania na słońcu na twarzach dorosłych szympansów pojawiają się ciemne plamy lub ciemnieją na całym ciele. Na podstawie skąpstwa zakładamy, że nasz ostatni wspólny przodek z szympansami również miał bladą skórę i ciemne futro. Bez względu na to, jak i kiedy nastąpiła drastyczna redukcja owłosienia ciała u ludzi (prawdopodobnie wraz z pojawieniem się rodzaju Homo w wieku około 2 milionów lat temu), jest wysoce prawdopodobne, że ciemnopigmentowana skóra przeszła bardzo szybką, bardzo silną selekcję pozytywną, gdy homininy straciły futrzaną sierść . Blada naga skóra stanowi wielkie zagrożenie dla zdrowia pod równikowym afrykańskim słońcem, dlatego zdecydowanie preferowana byłaby skóra o ciemnej pigmentacji. Dlaczego ciemno pigmentowana skóra jest lepsza dla ludzi w tropikach? Melanina, substancja nadająca kolor skórze, reguluje przenikanie szkodliwego promieniowania ultrafioletowego (UVR) ze słońca. Im więcej melaniny w skórze, tym bardziej może działać jako naturalny filtr przeciwsłoneczny, ponieważ w tropikach jest więcej promieniowania słonecznego niż gdziekolwiek indziej na świecie. Ryzyko oparzeń słonecznych i raka skóry jest znacznie zmniejszone u osób z ciemną pigmentacją. Jeden rodzaj UVR, UVA, uszkadza komórki i DNA. Powoduje rozpad kwasu foliowego (formy kwasu foliowego), opóźniając replikację DNA i podział komórek w zarodkach, co może następnie prowadzić do ich spontanicznej, naturalnej aborcji. Jest to bezpośredni wpływ UVR na kondycję u ludzi, ponieważ negatywnie wpływa na udaną reprodukcję. Skoro ciemna skóra jest korzystna, dlaczego niektórzy ludzie mają jasną skórę? W regionach położonych dalej od słonecznych tropików skóra o ciemnym zabarwieniu jest w rzeczywistości nieprzystosowana, ponieważ zbyt skutecznie blokuje promieniowanie UVB (inny rodzaj promieniowania UV). Penetracja UVB przez skórę jest konieczna, aby ludzie mogli metabolizować witaminę D w skórze. Na dużych szerokościach geograficznych z dala od równika występuje znacznie mniej promieniowania UVB. Mniej pigmentu w skórze pozwala na penetrację większej ilości promieniowania UVB i umożliwia niezbędny metabolizm witaminy D (która jest niezbędna, aby organizm między innymi wchłaniał wapń). W regionach, w których w atmosferze jest znacznie mniej promieniowania UVB, ludzie utracili znaczną część pigmentacji skóry pod presją selekcyjną ułatwiającą ten proces. Konsekwencje niedoboru witaminy D obejmują wystąpienie krzywicy, w której kości nie tworzą się prawidłowo, często uniemożliwiając choremu samodzielne poruszanie się. Kości nóg z krzywicą dosłownie wyginają się pod ich ciężarem. Co więcej, kobiety, które cierpią poważnie z powodu krzywicy mają zniekształcone kości miednicy, które utrudniają lub uniemożliwiają naturalny poród. To bezpośredni związek z fitnessem. W rzeczywistości jest bardzo niewiele obszarów poza tropikami, gdzie ludzie o ciemnej skórze mogą żyć przez cały rok i wchłonąć wystarczającą ilość promieniowania UVB. Wiele osób pochodzenia afrykańskiego mieszkających na przykład w Wielkiej Brytanii musi uzupełniać swoją dietę w żywność bogatą w witaminę D, taką jak ryby, aby żyć tam zdrowo. Analiza zegara molekularnego genu MC1R związanego z pigmentacją skóry wskazuje, że pojawienie się ciemnej skóry wynosi 1,7 miliona lat temu. Jest całkiem możliwe, że mieszkańcy Indonezji, Australii i południowego Pacyfiku wyewoluowali ciemną skórę oddzielnie, od przodków azjatyckich, które znacznie zmniejszyły ich pigmentację. Możliwe też, że jasna skóra wyewoluowała oddzielnie w Europie i Azji. Genetycy wkrótce będą mieli odpowiedzi na te pytania.
Początki Człowieków (43)
MITOCHONDRIALNA EWA I Y-CHROMOSOM ADAMA
Genetycy prześledzili podobieństwa w naszym mitochondrialnym DNA do jednej wspólnej matki nas wszystkich, a także ustalili hipotetycznego ojca wszystkich mężczyzn na Ziemi, śledząc ich odmianę chromosomu Y. Większość ludzi słyszała o mitochondrialnym DNA tylko z powodu odczucia "mitochondrialnej Ewy" wywołanej w latach 90. XX wieku. mtDNA to nie to samo co jądrowe DNA, które znajduje się w jądrze komórki. MtDNA znajduje się poza jądrem w organellach wytwarzających energię zwanych mitochondriami. Z kilku powodów mtDNA jest doskonałym źródłem informacji ewolucyjnych. Przede wszystkim genom dla mtDNA jest mniejszy niż jądrowy DNA i dlatego łatwiej go analizować niż jądrowy DNA. Po drugie, każda komórka ma tysiące więcej kopii mtDNA niż kopii jądrowego DNA, więc łatwiej jest z nią pracować w laboratorium. Po trzecie, mtDNA nie kumuluje mutacji tak szybko jak jądrowe DNA. I wreszcie, mtDNA jest dziedziczone tylko po matce, co oznacza, że każdy otrzymuje swoje mtDNA od matki. Nie ma rekombinacji z mtDNA ojca, ponieważ mtDNA w nasieniu ulega zniszczeniu w czasie zapłodnienia komórki jajowej. Ze względu na dziedziczne właściwości mtDNA, możemy prześledzić geny każdej żyjącej osoby wzdłuż linii matki, aż do samotnej matki wszystkich współczesnych ludzkich mtDNA. Ten przodek jest znany jako "mitochondrialna Ewa". Rebecca Cann i jej koledzy jako pierwsi oszacowali wspólnego przodka mtDNA dla wszystkich żyjących ludzi. Od tego czasu inni próbowali odpowiedzieć na to samo pytanie i zgłaszali różne, ale ogólnie podobne wyniki. Na podstawie porównania sekwencji między różnymi populacjami na całym świecie doszli do wniosku, że mamy wspólnego przodka mtDNA, który żył około 200 Kya w Afryce Subsaharyjskiej. Spośród populacji na całym świecie próbki afrykańskie wykazują największą zmienność w genomie mtDNA. Innymi słowy, populacje poza Afryką wykazują tylko podzbiór zmienności obserwowanej w próbie afrykańskiej. Zgodnie z teorią zegara molekularnego sekwencja o największej zmienności spowodowanej mutacjami miała najdłuższy czas na ewolucję lub akumulację tych mutacji. Analiza MtDNA potwierdza, że ludzie ewoluowali w Afryce przez dłuższy okres historii ewolucji człowieka niż gdziekolwiek indziej. Oczywiście "Ewa" nie była wówczas jedyną żyjącą samicą homininek. Inne kobiety żyły i rozmnażały się. Szczególnym punktem zainteresowania naszej Ewy jest to, że jest jedyną kobietą, której cała potomna linia miała przeżywające i rozmnażające się samice w każdym pokoleniu. Wszystkie inne linie, które wlokły się 200 000 lat temu, od tego czasu się skończyły i nie wniosły nic do naszego mtDNA. Porównując ogólnoświatową zmienność chromosomu Y, genetycy byli w stanie potwierdzić hipotezę mitochondrialnej Ewy. Ponieważ nie ma żeńskiego odpowiednika chromosomu Y, nie ulega on, podobnie jak mtDNA, rekombinacji. Tak więc chromosom Y jest przekazywany z ojca na syna. Wszystkie ludzkie chromosomy Y na świecie zbiegają się w "chromosomie Y Adama", który żył około 100 Kya. I podobnie jak mtDNA, ponieważ afrykańskie chromosomy Y wykazują największą zmienność, wynika z tego, że te rodowody ewoluowały najdłużej i że tam powstał ludzki chromosom Y. Niedawny wiek naszych wspólnych przodków, wynoszący 200 Kya i 100 Kya, jest mocnym dowodem przeciwko wieloregionalnemu modelowi pochodzenia człowieka i wspiera model Poza Afryką, który jest również nazywany hipotezą Ogrodu Edenu. Mitochondrialna Ewa i Y -chromosom Adama nie powinni być rozumiani dosłownie tak, jakby byli biblijną parą. Według ścisłej interpretacji Biblii Adam i Ewa byli pierwszymi ludźmi na ziemi. Jednak mitochondrialne Ewa i chromosom Y Adama są zgodne z teorią ewolucyjną, która zakłada, że mieli przodków i rówieśników. Są po prostu jedyną kobietą i jedynym mężczyzną, którego potomkowie przetrwali do dziś. Należy pamiętać, że chromosom Y Adama i mitochondrialna Ewa nie musieli żyć w tym samym czasie. Każdy z nich oznacza korzenie różnych aspektów naszej biologii, na których ewolucja budowała się od eonów. Pula genów chromosomów mtDNA i Y w Afryce zawiera więcej zmienności niż gdziekolwiek indziej, a zmienność genetyczna poza Afryką stanowi tylko podzbiór tych występujących na kontynencie afrykańskim. Dlatego z genetycznego punktu widzenia wszyscy ludzie są Afrykanami. Ze względu na ich tropikalne środowisko, w którym doświadczyliby wysokiego poziomu promieniowania UV, "Adam" i "Ewa" prawdopodobnie mieli ciemno zabarwioną skórę.
Wąskie gardło
Badania MtDNA i chromosomu Y, a także badania genów jądrowych wspierają ekspansję populacji z Afryki począwszy od 100 Kya. Poprzez statystyki i symulacje komputerowe oparte na analizach sekwencji różnych populacji na całym świecie genetycy z wielu różnych laboratoriów doszli do wniosku, że Homo sapiens doświadczył wąskiego gardła między 150 a 50 Kya. W tym momencie zmienność u ludzi była szczególnie niska. Badania nad zmiennością genetyczną w obrębie populacji i między populacjami sugerują, że w pewnym momencie globalna populacja ludzi mogła wynosić zaledwie 10 000, co byłoby zagrożonym poziomem, na równi z dzisiejszymi bonobo i gorylami górskimi. Od czasu utraty różnorodności genotypowej w wąskim gardle, ludzie doświadczyli zróżnicowania fenotypowego i rozwinęli zmienność regionalną, aby uczynić nas gatunkiem wielotypowym lub zmiennym, jakim jesteśmy dzisiaj.
Wrzody
Genetyczny korzeń wszystkich szczepów bakterii jelitowych człowieka (Helicobacter pylori) znajduje się w Afryce. Mniej więcej co drugi człowiek jest nosicielem H. pylori, czyli bakterii odpowiedzialnej za bóle żołądka i wrzody, i jest jedynym znanym mikroorganizmem, który może przetrwać w tak silnie kwaśnym środowisku. Na podstawie różnic genetycznych między populacyjnymi szczepami patogenu szacuje się, że początkowa infekcja H. pylori u ludzi wystąpiła około 60 Kya. Ponieważ sekwencje afrykańskich szczepów bakterii są bardziej zróżnicowane, przyjmuje się, że ewolucja musiała działać dłużej w Afryce, stąd też znajduje się tam korzeń przodków. Data 60 Kya sugeruje późniejszy scenariusz "poza Afryką" niż ten skonstruowany przez dowody na mitochondrialną Ewę, ponieważ ludzie wywieźli ze sobą H. pylori z Afryki. Filogenetyka molekularna pokazuje również, że H. pylori jest dalekim krewnym podobnych gatunków bakterii powodujących wrzody u lwów, tygrysów i gepardów. Genomy tych drobnoustrojów pokazują, że niektóre geny w postaci ludzkiej nadal funkcjonują, podczas gdy ich odpowiedniki w postaci kociej (Helicobacter acinonychis) już nie. Ponieważ jest mało prawdopodobne (tj. nie oszczędne), aby te geny zostały utracone, a następnie przywrócone w ludzkiej postaci, prawdopodobne jest, że ludzka postać, a nie forma wielkiego kota, jest bardziej podobna do drobnoustroju przodków. Dlatego jest całkiem możliwe, że bakteria została przeniesiona z ludzi na duże koty i, w oparciu o szacunki zegara molekularnego sekwencji dwóch gatunków, transmisja nastąpiła 200 Kya. Wielkie koty mogły zarazić się drobnoustrojem po zjedzeniu człowieka, a następnie przenosić go na inne koty w taki sposób, w jaki ludzie przenoszą go między sobą poprzez bliski kontakt fizyczny.
Początki Człowieków (42)
LUDZIE I SZYMPANSY: WĄSKI PRZEDZIAŁ
Odkąd Emile Zuckerkandl i Morris Goodman niezależnie porównali białka krwi afrykańskich małp i ludzi i ustalili, że są one nie do odróżnienia, niezwykłe podobieństwa molekularne między szympansami a ludźmi stały się oczywiste. Ale teraz, dzięki sekwencjonowaniu i mapowaniu (wskazaniu lokalizacji genów na chromosomach) całych genomów szympansów i ludzi, możliwe jest zidentyfikowanie genów odpowiedzialnych za ekspresję podobieństw, takich jak te białka krwi, a także różnic, takich jak wzory włosów i nawet rozmiar mózgu. Oprócz ludzi i szympansów, zsekwencjonowano wiele genomów, na przykład myszy, E.coli, rozdymki, pszczoły miodnej, ćmy jedwabistej i ryżu. W budowie są goryle, orangutany, bonobo i rezusy. Jak dotąd, jak można było przewidzieć, zwierzęta, które wyglądają podobnie, mają podobne genomy. Być może najbardziej upokarzającym odkryciem z mapowania ludzkiego genomu jest to, że ludzie mają od 20 do 25 000 genów, czyli tylko około dwa razy więcej niż u robaka lub muszki owocowej. Oczywiście, jeśli chodzi o badanie pochodzenia człowieka, najbardziej interesuje nas to, jak do siebie pasują ludzie i szympansy. Porównania sekwencji konsekwentnie wykazały, że ludzie i szympansy są najbliżej spokrewnionymi człekokształtnymi, dzielącymi około 99 procent ich kodu genetycznego. Genomy jednoznacznie dowodzą, że szympansy są naszymi najbliższymi żyjącymi krewnymi, tak jak przewidywał Darwin i tak samo jak wskazywały liczne inne badania, które doprowadziły do analiz całego genomu. W zależności od tego, jak spojrzy się na genomy ludzi i szympansów, można oszacować, że różnica w DNA jest większa niż 1 procent różnicy. Niektóre szacunki wskazują na różnicę do 8 procent. Różnice prostych par zasad nukleotydów są bardzo małe, rzędu mniej niż 2 procent różnicy, ale jeśli soczewka jest cofnięta i całe geny (segmenty par zasad) lub rodziny genów (geny, które dziedziczą się razem lub które współpracują ze sobą) są brane pod uwagę, wtedy różnice genetyczne między człowiekiem a szympansem mogą być większe. Ma to sens, ponieważ większość surowców jest taka sama dla szympansów i ludzi, po prostu jest inaczej modyfikowana w każdym z nich. Oboje mają włosy, ale po prostu inaczej ułożone i ułożone. Obie mają ręce i nogi, ale mają tylko różne długości. Obaj mają większe niż normalne mózgi i większą niż przeciętna inteligencję, ale ludzie są po prostu bardziej przesadni. Przy prawie żadnych różnicach sekwencji w porównaniu z szympansami ludzie nie są tak wyprowadzeni, jak wcześniej zakładano, i jeszcze wygodniej pasujemy do Drzewa Życia. Ale ludzie wyraźnie różnią się od szympansów, więc co sprawia, że człowiek nie jest szympansem i odwrotnie, co sprawia, że szympans nie jest człowiekiem? Czy możemy znaleźć geny odpowiadające za duży mózg, język, rozmiar zębów itp.? Naukowcy porównują i będą nadal porównywać genomy szympansa i człowieka, aby znaleźć regiony, które się różnią. Na przykład podczas skanowania sekwencji na komputerze naukowiec może znaleźć szczególnie interesujący obszar, który oddziela szympansy od ludzi. Następnie przejrzą bazy danych zmapowanych genów innych zwierząt, aby sprawdzić, czy te geny zostały zidentyfikowane i czy znane są ich funkcje. Jeśli nie, mogą znokautować gen ("gen kandydujący") u myszy, aby zobaczyć, jakie efekty, jeśli w ogóle, są widoczne w fenotypie z powodu utraty funkcji genu. Taki eksperyment na zwierzęciu modelowym pomaga określić funkcję genów, ponieważ myszy mają bardzo podobne genomy do szympansów i ludzi. Spośród wszystkich chromosomów, chromosom Y wydaje się nosić wiele różnic między szympansami a ludźmi. Około 30 procent naszych białek jest takich samych, a te, które różnią się od siebie, dzieli tylko kilka zmian aminokwasów. Wiele typów genów, które przeszły większą ewolucję u ludzi niż u szympansów - co jest śledzone przez zmiany sekwencji liczb w porównaniu z prymitywnymi genomami, takimi jak myszy - jest zaangażowanych w odporność, produkcję plemników i komórek jajowych oraz percepcję sensoryczną, taką jak węch. Porównania między genomem szympansa a ludzkim nic nie znaczą bez grupy obcej, która pomoże rozszyfrować, jakie cechy są prymitywne (wspólne z grupą obcą) i jakie cechy pochodzą (unikalne dla gatunku). Gdy genomy bonobo, goryla, orangutana i małpy rezus będą kompletne, będzie można stwierdzić, które geny powstały w linii szympansów, a które w linii ludzkiej. Mając zmapowane genomy blisko spokrewnionych grup obcych poprowadzą naukowców w kierunku genów, które odróżniają ludzi od innych małp człekokształtnych. Praktyczne i obiecujące aspekty sekwencjonowania całego genomu obejmuje znajdowanie leków na choroby i zaburzenia genetyczne, ale sekwencjonowanie całych genomów znacznie poprawia nasze zrozumienie relacji między genotypami i fenotypami oraz skomplikowanej biologii dziedziczenia. Przeglądanie genomów, zwłaszcza genów regulatorowych, pokazało nam, że dziedziczenie Mendla - biologia napędzana pojedynczymi genami i odrębnymi cechami - jest raczej wyjątkiem niż regułą. W rzeczywistości wiele cech jest poligenicznych, gdzie różne allele na więcej niż jednym genie, często na różnych chromosomach, są odpowiedzialne za jedną cechę. Ponadto cechy częściej są ciągłe niż dyskretne, na przykład w różnych odcieniach kolorów, w przeciwieństwie do po prostu czerni lub bieli. Efekty addytywne są powszechne, w którym to przypadku suma produktów każdego zaangażowanego genu daje fenotyp. Wydaje się, że większość różnic między DNA ludzi i szympansów ma związek z poziomem aktywności genów, a nie z samymi genami. Tak więc geny regulacyjne, które same regulują ekspresję lub wyrażają białka regulujące aktywność innych genów, są miejscem, w którym występuje zmienność człowiek-szympans. Dobrym wyjaśnieniem tego zjawiska może być to, że włączanie lub wyłączanie genu lub zmiana poziomów wyrażanych białek jest ewolucyjnie łatwiejsza niż zmiana samych genów, aminokwasów lub samych białek. Intrygująca różnica między szympansami a ludźmi występuje w przypadku genu MYH16. Szacuje się, że mutacja, która wystąpiła około 2,4 miliona milionów lat temu, sprawiła, że jest nieaktywna u ludzi, ale nadal dobrze funkcjonuje u szympansów, rezusów (Macaca mulatta) i innych naczelnych. MYH16 bierze udział w rozwoju mięśni szczęk do żucia, więc naukowcy stawiają hipotezę, że utrata funkcji tego genu jest związana z drastyczną zmianą morfologii czaszki u wczesnych członków rodzaju Homo około 2 milionów lat temu. Homininy w tym okresie przejściowym wyraźnie miały mniejsze zęby i mięśnie żucia i były w trakcie przyjmowania mięsożernego stylu życia, w którym kamienne narzędzia zaczęły zdejmować większość funkcjonalnego obciążenia z zębów i szczęk. Chociaż MYH16 prawdopodobnie nie jest jedynym genem zaangażowanym w kompleks mięśni żucia (ponieważ nie zniknął całkowicie wraz z mutacją), to z pewnością może ujawnić istotne wydarzenie w naszej ewolucji. Mapowanie całych genomów ujawniło, jak trudno jest znaleźć "gen dla" większości cech, nie tylko dlatego, że fenotypy są wyrażane przez skomplikowane orkiestracje genów, ale także dlatego, że mogą być pod silnym wpływem środowiska podczas rozwoju i podczas życia. Szukając określonych genów, genetycy mogą kontrolować czynniki środowiskowe, obserwując bliźnięta jednojajowe, które mają zasadniczo identyczne genotypy, ale dorastały w różnych środowiskach (tj. oddzielnych ciałach). Niestety takie eksperymenty są niemożliwe do przeprowadzenia z Homo erectus, ale zrozumienie wpływu środowiska na rozwój cech może pomóc paleontologom rozszyfrować, które cechy są ważne dla identyfikacji gatunków i śledzenia ewolucji, a które cechy są bezużyteczne do tych celów, ponieważ można je łatwo zmienić przez odżywianie lub inne czynniki niegenetyczne, środowiskowe. Ważny obszar badań genetycznych wykorzystuje loci cech ilościowych lub QTL w celu określenia, które cechy szkieletu i zębów skamieniałości są najbardziej przydatne do zrozumienia historii ewolucyjnej, w porównaniu z tymi, które są zbyt łatwo zmieniane przez środowisko podczas życia. Nieliczne różnice między ludźmi a małpami człekokształtnymi są w rzeczywistości genetyczną podstawą naszej szybkiej ewolucji kulturowej i ekspansji geograficznej po 200 Kya. Tylko kilka zmian genetycznych było koniecznych, aby prehistoria i historia ludzkości były możliwe.
"ŚMIECIOWE" DNA
Funkcja ponad 95 procent naszego DNA wciąż pozostaje tajemnicą. Oznacza to, że opisaliśmy kod, ale odkryliśmy, że większość z nich nie koduje białek. Geny mogą być oddzielone ogromną pustynią niekodującego DNA, które jest czasami nazywane "śmieciowym" DNA. Ale czy to bezużyteczne? Prawdopodobnie nie, ponieważ wśród sekwencji niekodujących znajdują się kluczowe regiony promotorowe, które kontrolują włączanie lub wyłączanie genów. Ludzki genom ma więcej niekodującego DNA niż jakiekolwiek inne zwierzę znane do tej pory i nie jest jasne, dlaczego. Co najmniej połowa sekwencji niekodującej składa się z rozpoznawalnych powtarzających się sekwencji, z których część została w przeszłości wstawiona przez wirusy. Te powtórzenia mogą zapewnić pewną swobodę ruchów genomowych. Oznacza to, że długie odcinki niekodującego DNA stanowią plac zabaw dla ewolucji. Posiadanie całego tego surowca dostępnego do mutacji i modyfikacji istniejących cech i zachowań lub jednoczesnego wyrażania nowych może być ogromną zaletą selekcyjną. Ludzie charakteryzują się zdolnością do elastyczności i szybkiej adaptacji, więc nasze śmieciowe DNA jest potencjalnie bezcennym wkładem w nasze człowieczeństwo.
Początki Człowieków (41)
Nowoczesne dowody
ZEGARY MOLEKULARNE
DNA to nie tylko podstawowy budulec życia; jest także rejestratorem danych. Sekwencja genetyczna osobnika opowiada o jego geograficznym pochodzeniu, a kod genetyczny gatunku opowiada o jego ewolucji. Porównując sekwencje DNA różnych organizmów, można określić, jak blisko spokrewnione są te dwa organizmy, a także można oszacować czas od ich wspólnego przodka. Podobieństwa w DNA zmniejszają się wraz ze spadkiem pokrewieństwa na poziomie indywidualnym, na poziomie gatunku i tak dalej. Identyczne bliźnięta rodzą się z identycznymi genomami. Normalne rodzeństwo ssaków dzieli średnio 50% swojego DNA, tak jak potomstwo dzieli średnio 50% swojego DNA z rodzicami. Wnuki dzielą 25 procent swojego DNA z dziadkami. I tak dalej. Niezależnie od tego, czy dwie osoby miały wspólną prapraprababkę 100 lat temu, czy też miały wspólny korzeń przodków 100 milionów lat temu, wszystko to jest zapisane w ich DNA. Metoda porównywania dwóch sekwencji, a następnie szacowania czasu od ich wspólnego pochodzenia, opiera się na zasadzie znanej jako "zegar molekularny". Pod koniec lat 60. Vincent Sarich i Allan Wilson byli pionierami w zastosowaniu tej koncepcji do ewolucji człowieka. Ich wstępna analiza porównująca afrykańskie małpy i ludzi zaowocowała drzewem z ludźmi i afrykańskimi małpami zgrupowanymi razem, z wyłączeniem małp azjatyckich. Ten wynik, wraz z potwierdzeniem wielu innych laboratoriów, przeczył założeniom opartym na anatomii i zachowaniu wielu naukowców przez większą część pierwszej połowy XX wieku. Długotrwały pogląd łączył małpy azjatyckie i afrykańskie z wyłączeniem ludzi, ale te nowe porównania genetyczne wykazały, że ludzie byli bliżej spokrewnieni z gorylami i szympansami niż małpy afrykańskie z małpami azjatyckimi. Szacowane daty rozszczepienia u małp uległy również zmianie po wynalezieniu zegarów molekularnych. Sarich i Wilson umieścili szympansa - człowieka, który podzielił się na około 5 milionów lat temu i od tego czasu liczne badania zegara molekularnego dały podobne szacunki między 8 a 4 milionami lat temu. Przed zegarami molekularnymi zakładano, że czas podziału był znacznie głębszy w prehistorii, między 30 a 20 milionów lat temu, co skłoniło niektórych wcześniejszych paleoantropologów do poszukiwania skamieniałości naszych dwunożnych przodków w znacznie zbyt starych skałach. Zasadniczo odległość genetyczną między dwiema próbkami ocenia się przez bezpośrednie porównanie nukleotydów w równoważnych pozycjach w każdej z dwóch sekwencji i zliczanie, ile razy się różnią. Metody zegara molekularnego można zastosować do genów, rodzin genów i, teoretycznie, do całych genomów. Są one również powszechnie stosowane do sekwencji DNA znajdujących się w mitochondriach, tzw. elektrowniach komórki, ponieważ wytwarzają energię. Chociaż DNA w jądrze komórkowym można wykorzystać do analizy zegara molekularnego, często preferowane jest DNA mitochondrialne (mtDNA), ponieważ mutacje w mtDNA kumulują się losowo i często z niewielkim wpływem na sprawność organizmu (chyba że wpływają na prawidłowe funkcjonowanie mitochondriów, prowadzące do zaburzeń mięśni). Neutralne regiony niekodujące są preferowane do stosowania w zegarach molekularnych, ponieważ bez działającej na nie selekcji mutacje w tych regionach gromadzą się ze stałą, podobną do zegara szybkością na pokolenie. Za pomocą zegarów molekularnych oszacowano i zweryfikowano czasy podziału wszystkich głównych grup naczelnych. Szacunki zazwyczaj obejmują pewien zakres czasu, ale mają tendencję do uśredniania mniej więcej tyle samo. Na zmienność szacowanych czasów podziału może wpływać zastosowana metoda kalibracji lub, jak omówiono powyżej, niezależnie od tego, czy stosuje się sekwencje neutralne. Zegar jest kalibrowany przy użyciu skamieniałości, co pomaga określić czas rozbieżności przy każdym kolejnym rozgałęzieniu drzewa. Dobrym punktem kalibracyjnym dla analizy zegara molekularnego czasów rozszczepienia małp i ludzi są dowody kopalne na rozbieżność między małpami Starego Świata a człekokształtnymi na granicy oligocenu i miocenu, 23 miliony lat temu.
Hybrydyzacja DNA-DNA
Jeden ze sposobów porównania sekwencji genetycznych jest następujący. DNA jest podgrzewane, aby zerwać wiązania między parami zasad, które utrzymują razem dwie nici. Dwuniciowa cząsteczka DNA dzieli się na dwie oddzielne nici w procesie zwanym "denaturacją". Następnie do mieszanki dodaje się zdenaturowane DNA porównywanych gatunków i całą próbkę pozostawia się do ostygnięcia. Podczas chłodzenia nici DNA łączą się ponownie ("anneal" lub "hybrydyzacja"), tworząc nowe kompletne, dwuniciowe cząsteczki DNA. Im bliżej gatunku są, filogenetycznie, im bardziej podobne będą ich sekwencje genetyczne (pary zasad), tym bardziej prawdopodobne jest, że ich nici DNA połączą się podczas tego procesu i tym ciaśniejsze będą ich wiązania. Aby ocenić, jak dobrze sekwencje do siebie pasują, są one ponownie podgrzewane. Jeśli denaturują się szybciej niż oryginalna cząsteczka DNA (z jednego gatunku), to wiązanie jest słabsze, ponieważ dopasowuje się mniej par zasad. Jeśli wymagają one równoważnej ilości czasu do rozszczepienia jak oryginalne DNA, to sekwencje są w zasadzie takie same
Początki Człowieków (40)
HOMO FLORESIENSIS
Jak widać na przykładzie plejstoceńskiej małpy Gigantopithecus, istota taka jak "Wielka Stopa" prawdopodobnie istniała w przeszłości, a skamieniałości z Indonezji sugerują, że "hobbici" też. W 2003 roku zespół australijskich i indonezyjskich naukowców kierowany przez Petera Browna przekopał dno jaskini zwanej Liang Bua i odkrył czaszkę z powiązanym szkieletem (LB 1) oraz szczątki szkieletowe co najmniej ośmiu innych małych osobników, które były nazywany "hobbitami". Do tej pory opublikowano tylko niektóre szczątki, ale są to obecnie najbardziej kontrowersyjne skamieniałości znane paleoantropologii. Te maleńkie homininy miały około 3 stóp wzrostu i miały mózgi wielkości szympansów. Opis fizyczny może brzmieć jak australopitek, ale w jaskini skojarzono z nimi złożone kamienne narzędzia i istnieją dowody na to, że polowali na słonie karłowate. Ponadto miejsce to datuje się na okres od 18 do 13 Kya, czyli znacznie później niż jakiekolwiek australopiteki, a morfologia zębów i czaszki przypomina wczesne Homo. Tak więc skamieniałościom nadano nowy gatunek, Homo floresiensis. Przypominając wczesne oceny neandertalczyków, krytycy twierdzą, że mała czaszka (380 cm3) jest patologiczna. Zakładają, że hominin miał stan podobny do małogłowie, w którym mózg nie rozwija się prawidłowo i pozostaje niebezpiecznie mały w wieku dorosłym. Zwolennicy nowego gatunku i jego "normalności" zwracają uwagę, że mikrogłowicy często mają zniekształconą czaszkę i twarz, a czaszka Liang Bua nie. W rzeczywistości odlanie LB1 pokazuje, że chociaż był mały, mózg H. floresiensis był wyrafinowany. Dwa różne geny (mikrocefalina i ASPM) po zmutowaniu mogą powodować małogłowie, ale niestety, nawet gdyby naukowcy byli w stanie wydobyć starożytne DNA z H. floresiensis, szanse na zachowanie sekwencji zawierającej którykolwiek z tych genów są bardzo małe. Chociaż do tej pory jest tylko jedna czaszka, istnieje kilka elementów pozaczaszkowych, które dzielą niewielki rozmiar ciała. H. floresiensis wygląda jak karłowaty gatunek hominina, chociaż nie dzieli kończyn z żyjącymi karłowatymi lub karłowatymi ludźmi. Krasnolud jest powszechny u zwierząt o dużych ciałach, takich jak słonie i hipopotamy, które ewoluują w izolacji na wyspach. Obecnie H. floresiensis jest postrzegany jako potomek indonezyjskiego H. erectus, który wyizolował się na Flores 800 Kya i poddał się presji ekologicznej życia na wyspie, zmniejszając swój rozmiar ciała. Potrzebne są dalsze badania, aby zrozumieć, w jaki sposób mózg ssaków, szczególnie mózg homininów, reaguje na karłowatość wyspową, aby jak najlepiej odpowiedzieć na małogłowie i hipotezy karłowatości, a także ustalić, czy H. floresiensis jest rzeczywiście potomkiem H. erectus.
Początki Człowieków (39)
ANATOMICZNIE NOWOCZEŚNI LUDZIE
Anatomicznie współczesny H. sapiens najpierw pojawił się w Afryce, a następnie rozprzestrzenił się na prawie każdy zakątek ziemi, zdolny do przetrwania we wszystkich klimatach i środowiskach. Drugie główne rozproszenie homininów, podjęte przez współczesnych ludzi, miało szersze rozproszenie niż początkowe rozproszenie przez H. erectus. Współcześni ludzie pojawiają się w Europie około 40 Kya, w Australii między 60 a 50 Kya, przebyli Cieśninę Beringa z Rosji do Ameryki Północnej około 30 Kya, a potem niemal natychmiast pojawiają się w Ameryce Południowej. Kilka szkieletów znaleziono w miejscu schronu skalnego Cro-Magnon w Les Eyzies we Francji, datowane na około 40 Kya. To z powodu tego znaczącego odkrycia Louisa Larteta w 1868 roku przyjęliśmy przydomek "Cro-Magnons" dla skamieniałości naszego gatunku. Do niedawna najstarsze współczesne skamieliny człowieka pochodziły ze stanowiska Herto w regionie Afar w Etiopii, datowane na około 160 Kya. Te trzy osobniki, w tym dwie czaszki, nadal zachowują małe mostki nadoczodołowe i inne prymitywne cechy, więc odkrywcy dali im podgatunek Homo sapiens idaltu, co oznacza "starszy". Jedna z czaszek miała ślady nacięć i została wypolerowana prawdopodobnie przez noszenie ze sobą po śmierci osobnika, co może wskazywać na wczesny etap rytualnego zachowania. Teraz, gdy miejsce to zostało przeredagowane do 195 Kya, najwcześniejsze skamieniałości w pełni współczesnych ludzi to dwa szkielety, Omo I i II, które znaleziono w pobliżu rzeki Omo w Etiopii, naprzeciw północnej granicy Kenii. Te skamieliny potwierdzają szacunki zegara molekularnego, które wskazują na pochodzenie naszego gatunku z Afryki Wschodniej na około 200 Kya. Chociaż istnieją różnice między współczesnymi populacjami i wewnątrz nich, wszyscy ludzie mają wspólne cechy czaszki, które odróżniają nas od wcześniejszych homininów. Te cechy obejmują wysokie, okrągłe czaszki, pionowe płaskie czoła z małymi lub żadnymi łukami brwiowymi, podbródki i średni rozmiar mózgu 1350 cm3 - średnio nieco mniejszy na rozmiar ciała niż neandertalczycy, ale większy niż jakikolwiek inny hominin. Zwężenie oczodołowe jest niewielkie lub nie ma go wcale, co oznacza, że mięśnie do żucia są ekstremalnie zredukowane, a mózg jest rozszerzony w ich miejsce. Mała szczęka ma wydatny podbródek i małe zęby. Znaczniki mięśni na czaszce i całym szkielecie są mniej mocne, a kości są bardziej smukłe lub smukłe i słabe niż poprzednie homininy. Twarz jest schowana pod czaszką i jest mniej prognatyczna niż u poprzednich homininów. Najwcześniejsi ludzie mieli również tropikalne proporcje ciała. Gdy pojawili się współcześni ludzie, niemal natychmiast wypełnili nisze tropikalne, umiarkowane i arktyczne. Pierwsi ludzie byli zbieraczami, zbieraczami, łowcami dużych i małych zdobyczy oraz rybakami. Podróżowali daleko, aby zdobyć surowce lub za nie handlować. Ich zachowanie i kulturę charakteryzuje złożoność, elastyczność i innowacyjność. Po raz pierwszy zrobiono z kości igły (najstarsze to 26 Kya), szydła do szycia odzieży i miotacze włóczni (lub "atlatle"), które dodają niesamowitej prędkości i odległości rzuconej włóczni. Koraliki, wisiorki i inne dowody osobistego ozdoby są wszędzie, zanim współcześni ludzie przejmą władzę w górnym paleolicie lub późnej epoce kamienia.
Początki Człowieków (38)
NARZĘDZIA KAMIENNE
Narzędzia kamienne stają się z czasem mniejsze, bardziej wyspecjalizowane i trudniejsze do wykonania. Najwcześniejszym, najbardziej prymitywnym przemysłem narzędzi kamiennych rozpoznanym w danych archeologicznych jest przemysł olduwajski lub tradycja wczesna epoka kamienia. Jest to kultura prostych, surowych rdzeni i ostrych płatków z zaledwie garstką nazwanych typów w zestawie narzędzi, takich jak siekacze, młoty i skrobaki. Płatki trzymane między kciukiem a palcem wskazującym mogą być używane jak skalpel do oskórowania i uboju zwłok zwierzęcych. Celowo wyprodukowane płatki różnią się od płatków wytworzonych przez Matkę Naturę bliznami powstałymi podczas produkcji: ze względu na klasyczny wzór pęknięcia, zwykle w miejscu, w którym młotek uderzył płatek z rdzenia i pod spodem, znajduje się wybrzuszenie lub "bulwa perkusja." Skrobaki i siekacze Olduwa to narzędzia wielościenne i kuliste które byłyby przydatne w przypadku szerokiej gamy przygotowywania posiłków, zarówno ze zwierząt, jak i roślin. Młoty nie tylko wybijają płatki z miękkich skał, ale także rozbijają orzechy i kości w celu uzyskania pożywnego szpiku. Pierwsze narzędzia kamienne zostały wyprodukowane około 2,6 mln lat temu, a dowody pochodzą z miejsca Kada Gona w regionie Hadar w Etiopii. Inne wczesne miejsca to Omo i Bouri w Etiopii oraz wąwóz Olduvai w Tanzanii. Jakie gatunki homininów wytwarzały narzędzia Olduwa? Pewne jest, że robił je wczesny Homo, ale czy to oni je wymyślili, czy zrobili australopity? I czy zrobił je Paranthropus? Oczywistą zaletą narzędzi kamiennych byłoby użycie ich do rzeźni kości zwierząt na mięso i rozbijanie kości w celu uzyskania bogatego w składniki odżywcze szpiku w środku. Zwykle zakładamy, że dopiero wczesny Homo jadł mięso. Ale byłyby również przydatne dla australopitów i parantropów do rozłupywania orzechów oraz do kopania bulw i innych podziemnych pokarmów. Jak dotąd nadal nie ma jednoznacznych dowodów, aby wiedzieć, czy jest to jeden czy inny sposób. Opierając się na prostej technologii Olduwańskiej, przemysł aszelski, który powstał około 1,6 miliona lat temu, zawierał bardziej wyrafinowane narzędzia, w tym siekiery ręczne i kilofy. Te narzędzia były nadal prymitywne, ale wyglądały bardziej symetrycznie niż poprzednie. Nazywa się je "dwójkami", ponieważ kamień jest łuszczony i formowany z obu stron, aby nadać mu symetryczny wygląd. W przeciwieństwie do większości przemysłu Olduwańskiego, nawet niespecjalista może wyraźnie przypuszczać, że wytwarzanie narzędzi aszelskich wymagało znacznych umiejętności. Tradycja aszelska, nazwana tak od miejsca jej pierwszego odkrycia we Francji, w niektórych miejscowościach trwała do 1,5 miliona lat. Oczywiście narzędzia z zestawu aszelskiego były przydatne dla H.erectus i ludzi archaicznych do przetwarzania żywności. Przez wiele lat uważano, że pojawienie się technologii aszelskiej pozwoliło H. erectus z powodzeniem promieniować poza Afrykę. Jednak inne zwierzęta, takie jak słonie, również pochodzą z Afryki i rozproszyły się po Starym Świecie bez pomocy kamiennych narzędzi. Co więcej, Dmanisi, najstarsze miejsce homininów poza Afryką, w którym zachowały się narzędzia, ma tylko narzędzia Olduwa. Na stanowiskach azjatyckich generalnie brakuje narzędzi aszelskich, co po raz pierwszy poważnie rozważał archeolog Hallum L. Movius. Narysował teoretyczną linię progową ("linia Moviusa") w całej Eurazji, poza którą na Wschodzie nie znaleziono żadnych narzędzi aszelskich. Istnieje wiele powodów, dla których Acheulean może być rzadkością w Azji i Indonezji. Jest całkiem możliwe, że homininy rozproszyły się z Afryki przed wynalezieniem technologii aszelskiej. Ponadto wschodnie homininy mogły w większym stopniu polegać na organicznych narzędziach wykonanych z takich rzeczy jak bambus, który się nie konserwował. Linia Moviusa może jednak nie trwać długo, ponieważ w 2000 roku archeolodzy poinformowali, że odkryli narzędzia przypominające toporki ręczne (prawie nie do odróżnienia od afrykańskich) w basenie Bose w Chinach datowane na 800 Kya. Zanim archaiczni ludzie pojawią się w środkowym paleolicie, wykazują poprawę w stosunku do Acheulean, która obejmuje użycie techniki Levallois, która jest metodą stosowaną do kontrolowania wielkości i kształtu płatków. Przygotowując najpierw rdzeń, z jednego rdzenia można wytworzyć więcej płatków, ostrzejszych płatków i dłuższych płatków. Jest to proces, który wymaga kilku kroków i wskazuje na wyższą inteligencję. Neandertalczykom w przeważającej części przypisuje się tworzenie narzędzi Mouster . Użyli Levalloisa i innych przygotowanych podstawowych technik. Narzędzia wykazują zwiększoną różnorodność, do wykonania których wymagane są miękkie młotki, takie jak kości i poroże, są one często retuszowane lub łuszczone i przekształcane w celu ich ostrzenia i honowania. Najwyraźniej do tworzenia narzędzi Mousterian potrzebny jest mózg, a nie siła. Niektórzy neandertalczycy przeszli do tworzenia bardziej zaawansowanych narzędzi Ch?atelperronian, które są bardzo podobne, jeśli nie identyczne, do niektórych współczesnych ludzkich zestawów narzędzi z tego samego okresu. Pojawienie się jakiegokolwiek konkretnego gatunku hominina nie koreluje z jednoczesnym pojawieniem się konkretnego przemysłu narzędzi kamiennych. Na przykład skamieniałości H. erectus pojawiają się przed narzędziami aszelskimi, a tak zwany rewolucyjny zestaw narzędzi współczesnego człowieka/górnego paleolitu pojawia się dopiero około 150 000 lat po pierwszych ludzkich skamieniałościach. Do 45 Kya ludzie wytwarzali ostrza (bardzo długie płatki) i mikrolity (małe płatki, które również były przymocowane do innych narzędzi). Najwcześniejsza kultura narzędziowa z górnego paleolitu, zwana oryniacką, zastąpiła wszędzie kultury neandertalskie o 28 kya. Podobnie jak inne aspekty ludzkiego zachowania, ludzie posunęli się do skrajności w wytwarzaniu narzędzi. Wynaleźli większość rodzajów narzędzi, wykorzystali różnorodne materiały (kamień, kości, rogi, poroże i zęby), pozyskiwali surowce z odległości nawet setek mil, a także wyprodukowali najmniejsze narzędzia w historii.
Początki Człowieków (37)
Neandertalczycy
Pierwszymi skamieniałościami homininów nie będących ludźmi znanymi nauce byli neandertalczycy (wymawiane Nee-an-der-tals) lub Homo neanderthalensis. Były to krępe, muskularne gatunki o baryłkowatej klatce piersiowej, znane z miejsc w Europie, na Bliskim Wschodzie i w Eurazji. Nazwa Neandertalczyk pochodzi od lokalizacji jednego z najwcześniejszych odkryć tego gatunku, w Dolinie Neander w Niemczech, w 1856 roku. W oparciu o błędną interpretację anatomii i chęć wywyższenia człowieka spośród naszych ewolucyjnych kuzynów, neandertalczyków początkowo uważano za małpy człekokształtne - podobni do dzikusów z małym intelektem i zgarbionym, wleczonym krokiem. Stereotypy przetrwały do dziś, pomimo wszystkich współczesnych dowodów, że jest inaczej. Neandertalczycy istnieli od około 350 do 28 Kya, a wiele stanowisk omawianych w ramach Archaików jest również uważanych za stanowiska neandertalskie. Pod koniec swojego istnienia neandertalczycy coraz bardziej różnili się wyglądem od ludzi współczesnych, coraz bardziej upodabniali się kulturowo do ludzi współczesnych (jest to kwestia dyskusyjna), a następnie zniknęli. "Klasyczni" neandertalczycy, najbardziej odmienne anatomicznie i stereotypowe okazy, występują w Europie. Czaszka ma niewątpliwie duże, zaokrąglone łuki brwiowe w kształcie litery M, które wystają nad duży otwór na nos. Środek twarzy wystaje na nos i wydaje się być wysunięty w porównaniu z resztą twarzy, która wydaje się odsuwać od nosa w kierunku kości policzkowych. Szczęka nie ma kości podbródka, jak u ludzi, a także ma dodatkową przestrzeń zwaną przerwą zatrzonową za trzecimi zębami trzonowymi. Trzonowce nazywane są "taurodontami" ze względu na ich mocny i nadęty lub "byczy" wygląd. Niska, płaska kość czołowa, długa puszka mózgowa i wybrzuszony kok potyliczny (tj. guzek) mają największy mózg - dla wszystkich homininów, w tym ludzi - o pojemności 1300-1600 cm3. Niewątpliwy jest również szkielet pozaczaszkowy neandertalczyków. Niezwykle mocne kości wskazują, że neandertalczycy byli silni, muskularni i wysportowani. Długie kości neandertalczyków są grube i zakrzywione z powodu nałożonych na nie ogromnych sił mięśniowych. Pierwotnie za te zakrzywione kości przypisywano krzywicę, chorobę związaną ze złym odżywianiem. Szkielety neandertalczyków często wykazują ślady urazu. Eric Trinkaus porównał ich szczególny schemat obrażeń do rodzajów złamanych i zagojonych kości, jakich doświadczają amerykańscy jeźdźcy rodeo. Neandertalczycy niekoniecznie jeździli na dzikich zwierzętach, ale prawdopodobnie wchodzili z nimi w niebezpiecznie bliski kontakt. Badania kości ramion przeprowadzone przez Stevena Churchilla sugerują nawet, że woleli oni wbijać włócznie w swoją ofiarę niż nimi rzucać, co potwierdza interpretację ich niebezpiecznych technik łowieckich. Zęby neandertalczyków pracowały tak samo ciężko jak ich ciała. Często ich siekacze są zużyte, a nawet mają na sobie ślady po kamiennych narzędziach, tak jakby neandertalczycy używali swoich szczęk jako imadła do przygotowywania jedzenia lub skór zwierzęcych. Pomimo surowego stylu życia, neandertalczycy dokonali postępów w materiałach kulturowych, które zbliżały się do współczesnych ludzi. W jednym z najnowszych (lub ostatnich) stanowisk neandertalskich w Grotte du Renne we Francji, datowanym na 34 Kya, zachowały się perforowane i rowkowane zęby, kość słoniowa i koraliki kostne, które prawdopodobnie były używane do osobistych ozdób. Jeśli tradycyjnie uważa się, że tylko współcześni ludzie zachowywali się w ten sposób, czy należy założyć, że neandertalczycy handlowali lub wchodzili w interakcje ze współczesnymi ludźmi? Dyskutuje się, czy neandertalczycy i współcześni ludzie wyewoluowali to zachowanie niezależnie lub czy kiedykolwiek weszli w interakcje. Krąg kości mamutów z paleniskami, narzędziami i kośćmi zwierzęcymi w miejscu na Ukrainie pokazuje, że neandertalczycy tworzyli obozy jak współcześni ludzie. Dlatego stricte "jaskiniarski" wizerunek neandertalczyków może być wynikiem zachowania zapewnianego przez jaskinie. Nie ma jeszcze igieł związanych z neandertalczykami, więc możliwe, że nie tworzyli oni ubrań. Nie wyklucza to jednak prawdopodobieństwa, że nosili skóry zwierzęce, ponieważ żyli w zimnym klimacie. Nie wiadomo, dlaczego neandertalczycy zniknęli około 28 Kya. Być może zbyt dobrze przystosowali się do zimna, aby nie mogli dłużej przetrwać między epokami lodowcowymi. Być może współcześni ludzie byli lepsi w wypełnianiu podobnej niszy i prześcigali ich. A może współcześni ludzie zalali pulę genów neandertalczyków, krzyżując się z nimi. Hipoteza autorstwa Stevena Kuhna i Mary Stiner sugeruje, że za upadek neandertalczyków można przypisać sposób ich polowania. Albo alternatywnie, sukces współczesnych ludzi może być częściowo spowodowany wprowadzeniem podziału pracy między płciami. Dowody wskazują, że całe populacje neandertalczyków brały udział w polowaniu na grubą zwierzynę. Jest to zupełne przeciwieństwo wczesnych współczesnych ludzi, którzy - wzorując się na żyjących łowcach-zbieraczach lub społecznościach zbieraczy, takich jak lud San z pustyni Kalahari w południowej Afryce - kazali mężczyznom polować na dużą zwierzynę, podczas gdy kobiety zbierały drobną zwierzynę i rośliny. Taki podział pracy zapewnia ciągłe i zróżnicowane zaopatrzenie w żywność, ponieważ pozyskiwanie mięsa od dużych zwierząt może być niejednolite i nieprzewidywalne. Neandertalczycy niebezpiecznie polowali z włóczniami (nie było jeszcze łuków i strzał ani nawet miotaczy włóczni) i wchodzili w bliski kontakt ze swoją ofiarą. Możliwe, że brały w nich udział kobiety i dzieci. Na stanowiskach neandertalskich nie ma igieł kostnych, bardzo niewiele szczątków małych zwierząt i kamieni mielących do przygotowywania pokarmów roślinnych. Są to artefakty związane z rolami kobiet w macierzystych społeczeństwach ludzkich i zostałyby pozostawione, gdyby kobiety i dzieci nie brały udziału w polowaniach. Strategia neandertalczyków mogła być ukierunkowana na bogate w składniki odżywcze źródła białka, ale mogła być zbyt wyspecjalizowana i mogła zagrażać ich rdzeniowi reprodukcyjnemu aż do śmierci gatunku.
Początki Człowieków (36)
ARCHAICZNY HOMO SAPIENS
Homininy, które żyły w całym Starym Świecie między 800 a 125 Kya, należą do kategorii zwanej "Archaiczny H. sapiens". Archaiki stanowią grupę przejściową między H. erectus a współczesnym człowiekiem i obejmują neandertalczyków, o których mowa osobno. Archaiki wciąż różniły się anatomicznie od współczesnych ludzi, głównie pod względem czaszek o grubych kościach i niskim sklepieniu, z wydatnymi łukami nadoczodołowymi, spadzistymi czołami i małymi podbródkami. Przypominające wcześniejsze H. erectus, ich czaszki zachowują długi i niski profil, ale wykazują zwiększoną krągłość, jak współcześni ludzie, z maksymalną szerokością większą niż otwory na uszy. Piłki sportowe są dobrą metaforą kontrastów kształtowych między czaszką gatunku (czaszka bez twarzy). H. erectus crania są jak futbol amerykański, archaiki są jak piłki do rugby, a współcześni ludzie są jak okrągłe piłki do piłki nożnej. Twarz archaiczna jest nadal prognatyczna, ale znacznie mniej niż wcześniejsze homininy. Ich zęby były nieco większe niż u współczesnych ludzi, chociaż były znacznie mniejsze niż zęby wcześniejszych homininów. Szkielety archaiczne są solidne ze względu na ciężką życiową muskulaturę, a ich rozmiar mózgu wynosił średnio około 1200 cm3 (w zakresie od 900 do 1400 cm3), czyli praktycznie tyle samo, co rozmiary współczesnego mózgu człowieka. Pomimo fizycznych podobieństw do współczesnego H. sapiens, brakowało im zdolności kulturowych, które wyróżniają nasz gatunek. Skamieniałości archaiczne znajdują się w połączeniu z narzędziami aszelskimi i bardziej złożonymi, ale nie najbardziej złożonymi, jakie wynaleźli współcześni ludzie. Nie jest pewne, którzy archaicy prowadzili do neandertalczyków czy współczesnych ludzi, ale pomimo niejednoznaczności ogólnej kategorii, istnieje kilka grup skamielin, które są uważane za ich własny gatunek. Homo heidelbergensis obejmuje większość archaików w Europie, basenie Morza Śródziemnego i Afryce, datowanych na okres od 600 do 125 Kya. Nazwa H. heidelbergensis została nadana żuchwie znalezionej w Heidelbergu w Niemczech w 1907 roku. Jednak najwcześniejszymi przedstawicielami tego gatunku są czaszki afrykańskie znalezione w Kabwe w Zambii i Bodo w Etiopii, datowane na 600 Kya i związane z artefaktami Acheulean. Archaiczne skamieliny z Azji pochodzą z miejsc takich jak Dali w Chinach, datowanych na około 200 Kya. Azjatyckie czaszki archaiczne mają pewne prymitywne cechy z H. erectus, ale mają większe możliwości czaszki i nie są prognatyczne. Homo heidelbergensis w Europie prawdopodobnie dał początek eandertalczykom. Stanowisko w Atapuerca w Hiszpanii, datowane na około 800 Kya, wyprodukowało kilka szkieletów przodka Homo, które są potencjalnymi, bardzo wczesnymi prekursorami neandertalczyków. Liczne szkielety zostały również odkryte w pobliskim miejscu Sima de los Huesos ("doł kości") datowane na późniejszy wiek 350 Kya i mogą, ale nie muszą, być potomkami wcześniejszych populacji lub przodkami neandertalczyków. Wcześniej w zapisie kopalnym czaszka z Petralony w Grecji, datowana na około 400 Kya, mogła być przodkiem neandertalczyków z ogromnymi nadczołami i dużym otworem nosowym. Jeszcze starsza czaszka z Arago we Francji, datowana na około 450 Kya, również przypomina neandertalczyka ze swoją dużą, stosunkowo wystającą twarzą. Archaicy są pierwszymi homininami, którzy mają powiązane dowody archeologiczne budowanych schronów czy to ze starych dołków posłupowych, czy też z zachowanych materiałów. W Lazaret Cave we Francji zbudowano schronienie przy ścianie jaskini, a w Terra Amata (również we Francji) istnieją dowody na sezonowe lub krótkotrwałe użytkowanie schronienia. Dzięki Archaikom znajdujemy pierwszy wyraźny dowód używania jaskini przez homininów. W tym momencie istnieje również wiele dowodów na polowanie na grubą zwierzynę. Na wysokości 250 Kya w miejscu La Cotte de St. Brelade na Wyspach Normandzkich między Wielką Brytanią a Francją znajdują się szkielety mamutów i nosorożców włochatych, związane z licznymi płatkami kamienia. Zwłoki zostały przetworzone do jedzenia, czaszki zostały rozcięte w celu wydobycia mózgów, a kości spalono. Liczne szkielety w kolekcji sugerują, że zostały zepchnięte z klifów, a ponieważ nie ma dowodów na istnienie palenisk długoterminowych, miejsce to było prawdopodobnie tymczasowym obozem myśliwskim. Najwcześniejsze zachowane włócznie pojawiają się również w tym czasie, około 400 Kya i pochodzą z Sch¨oningen w Niemczech. Zostały wykonane z wewnętrznej części pnia świerka i mają utwardzone w ogniu końcówki i zwężające się uchwyty. Równowaga włóczni sugeruje, że były ważone do rzucania, ale jest tak samo możliwe, że były pchnięte tylko podczas użycia. W miejscu włóczni znajdowały się również setki kości koni, które zostały zabite (prawdopodobnie za pomocą włóczni), a następnie wyrżnięte.
Początki Człowieków (35)
HOMO ERECTUS
Pierwsze odkryte skamieniałości H. erectus to jarmułka (górna część czaszki, która nie obejmuje twarzy ani dolnej części, w której znajduje się otwór magnum), trzonowiec i kość udowa z Trinil na Jawie w 1891 roku. Te znaleziska obejmują brakujące ogniwo, które Eugèene Dubois wyruszył z Holandii, aby znaleźć. Od tego czasu paleoantropolodzy zebrali okazy H. erectus z Etiopii, Kenii, Tanzanii, RPA, Maroka, Włoch, Indii, Chin ("Człowiek z Pekinu") i Indonezji ("Człowiek z Jawy" i "Człowiek z Solo"). Gatunek oznacza pierwsze rozproszenie homininów poza Afryką, co odpowiada poważnej zmianie w ekologii. Powszechnie przyjmuje się, że H. erectus wyewoluował w Afryce, ponieważ znajdują się tam homininy, które go poprzedzały. Jednak najwcześniejsze daty występowania H. erectus ze stanowisk w Gruzji i Indonezji wskazują na 1,8 miliona lat temu. Aby odwrócić afrykańskie pochodzenie gatunku, starsze skamieniałości H. habilis lub australopiteków musiałyby zostać znalezione poza Afryką, a jak dotąd tak się nie stało. H. erectus obejmuje prawie 2 miliony lat ewolucji homininów, przy czym ostatni bastion tego gatunku utrzymuje się w Indonezji do około 30 000 lat. H. erectus różni się od poprzednich homininów kilkoma cechami. Pojemność czaszki jest zwiększona w zakresie od około 750 cm3 do 1200 cm3, co mieści się w zakresie rozmiarów współczesnego ludzkiego mózgu. Czaszki H. erectus nie mają podbródków, cofniętych czoła i masywnych, krótkich, wystających twarzy, zwieńczonych dobrze rozwiniętymi naczyniami (które w rzeczywistości wyglądają bardziej jak pojedynczy poziomy wizjer w porównaniu z późniejszymi naczołami hominina, które są mniejsze i czasami mają kształt litery "m"). Mieli też mniejsze zęby i grubsze kości czaszki niż ich poprzednicy. Wraz z pojawieniem się H. erectus po raz pierwszy pojawiły się ludzkie rozmiary i proporcje. Szkielet był prawie nie do odróżnienia od współczesnego człowieka, chociaż był bardziej wytrzymały niż nasz. Samce i samice były bardziej wyrównane pod względem wielkości i kształtu ciała oraz zębów. Najbardziej kompletnym w historii szkieletem H. erectus jest chłopiec Nariokotome, czasami nazywany także chłopcem Turkana (KMN-WT 15000). Obie nazwy odnoszą się do regionu geograficznego, w którym wykopano szkielet - stanowiska Nariokotome po zachodniej stronie jeziora Turkana w Kenii. W 1985 roku Kamoya Kimeu odkrył chłopca, dostrzegając mały kawałek czaszki, który skłonił zespół wykopaliskowy do odkrycia całej czaszki tuż przy powierzchni, a następnie do reszty szkieletu głębiej w ziemi. Mikroskopijna struktura zębów chłopca Nariokotome wskazuje, że zmarł w wieku około ośmiu lat, a nie jedenastu czy dwunastu lat, jak wcześniej sądzono. W wieku ośmiu lat chłopiec Nariokotome miał 5 stóp 3 cale i urósłby do ponad 6 stóp, gdyby dożył dorosłości. Duża dziurawa blizna na jego żuchwie z ropnia korzenia zęba może bardzo dobrze wskazywać na przyczynę jego śmierci. Takie zmiany, jeśli nie są leczone, mogą prowadzić do infekcji, która może rozprzestrzenić się do krwioobiegu i zatruć organizm poprzez zagrażający życiu stan zwany posocznicą. Szkielet chłopca z Nariokotome wykazuje przystosowanie do w pełni zaangażowanego i wydajnego dwunożności bez trwałych cech nadrzewnych. Jego proporcje kończyn wskazują, że rządy Allena działały na ludzi już we wczesnym plejstocenie; jego długie dystalne segmenty kończyn i wąska miednica są podobne do współczesnych ludzi żyjących w gorącym klimacie tropikalnym. Czaszki azjatyckiego H. erectus mają wspólne elementy wyglądu. Mają jeszcze bardziej wyraziste łuki brwiowe, wyraźniejsze kile na całej czaszce i grubsze kości czaszki. Ta zmienność powoduje, że niektórzy badacze łączą wszystkie H. erectus z całego Starego Świata w jeden gatunek obejmujący od 1,8 Mya do 30 Kya. Ale ci, którzy podkreślają różnice w skamieniałościach indonezyjskich, zachowują te z H. erectus, ale skamieniałości z Afryki i Georgii nazywają Homo ergaster, co oznacza "człowiek pracujący".
Dmanisi
Położona u podnóża gór Kaukazu wioska Dmanisi w Gruzji początkowo zainteresowała archeologów ze względu na swoją średniowieczną fortecę z IX wieku. Na początku lat 80. w ścianach dołów magazynowych, które średniowieczni mieszkańcy wykopali w twierdzy, odkryto skamieniałości. Jedna ze skamieniałości została zidentyfikowana jako szczęka H. erectus. Od tego czasu w Dmanisi odkryto co najmniej cztery czaszki i jeden częściowy szkielet H. erectus, a także setki surowych narzędzi kamiennych. Warstwa zawierająca skamieliny w Dmanisi jest datowana na około 1,78 miliona lat temu, wiążąc ją z Trinil w Indonezji, jako najwcześniejsze miejsce homininów poza Afryką. Ze względu na stały przepływ skamieniałości homininów z tego miejsca, Dmanisi jest głównym miejscem do badania pierwszego rozprzestrzeniania się homininów z Afryki. Z nielicznych dotychczas odkrytych skamieniałości jasno wynika, że były to osobniki duże i małe. Biorąc pod uwagę zmienność wykazywaną przez najwcześniejsze Homo (habilis i rudolfensis), która nakłada się w czasie, być może taką zmiennością był normalny dymorfizm płciowy lub normalna zmienność populacji. Być może w Dmanisi istnieją dwa gatunki homininów. Kamienne narzędzia w Dmanisi to bardzo prymitywne elementy litu Olduwańskiego. Nie są to narzędzia aszelskie, które normalnie kojarzy się z H. erectus.
Początki Człowieków (34)
RODZAJ LUDZKI
W przypadku rodzaju Homo otrzymujemy najwcześniejsze dowody kopalne na rozprzestrzenienie się poza Afrykę. Jak w przypadku wszystkich początków linii, identyfikacja pierwszego przedstawiciela rodzaju Homo w zapisie kopalnym jest trudna. Istnieje kilka okazów z Etiopii, Kenii i Tanzanii, które są godnymi kandydatami. Problem tkwi w definicji Homo. Cechy, które uzasadniają włączenie do ekskluzywnego klubu rodzaju ludzkiego, nie są ogólnie uzgodnione. Nie jest już modne przypisywanie homininów Homo w oparciu o zdolność do wytwarzania narzędzi z kamienia, ponieważ trudno było jednoznacznie wykazać związek między najwcześniejszymi narzędziami a konkretnym gatunkiem. Geografia i wiek Homo, Paranthropus i Australopithecus w późnym pliocenie i wczesnym plejstocenie pokrywają się w znacznym stopniu, więc nie możemy definitywnie wykluczyć australopiteków lub Paranthropus z gildii kamiennych narzędzi. Inni badacze opierają się raczej na anatomicznych niż behawioralnych dowodach włączenia do rodzaju Homo. Większość badaczy uważa Homo habilis za członka rodzaju ludzkiego ze względu na różnice między nim a australopitekami, takie jak mniejsza i bardziej płaska twarz, mniej opadające czoło, bardziej zaokrąglona czaszka, mniejsze szczęki i zęby, cieńsze kości czaszki i brak grzebienie czaszki. Od dawna przestrzegany jest wymóg dotyczący wielkości mózgu: czaszki o pojemności czaszki powyżej 600 cm3 są bezpiecznie uważane za należące do rodzaju Homo. Dopóki czaszki z taką pojemnością czaszki nie mają cech australopiteków ani parantropów, zasada ta jest podtrzymywana i jak dotąd nie znaleziono żadnej, która by jej nie przeczyła. Opierając się na proporcjach kończyn przypominających małpy, zrekonstruowanych z fragmentarycznych szczątków szkieletowych i na podstawie wielu podobieństw do australopitów w przeciwieństwie do późniejszego H. erectus, H. habilis jest coraz częściej umieszczany w rodzaju Australopithecus. Jednym z powodów tej zmiany taksonomicznej jest częściowy szkielet OH 62, H. habilis z wąwozu Olduvai. Rozdrobnione szczątki tej starej kobiety zostały znalezione w pobliżu polnej drogi po tym, jak zostały przypadkowo przejechane. Po zrekonstruowaniu kończyn i oszacowaniu proporcji ciało OH62 jest bardzo małpopodobne, czasem nawet bardziej małpopodobne niż niektóre australopity. Niektóre skamieniałości, jak ta w szczególności, przysparzają więcej kłopotów niż dostarczają odpowiedzi. Dopóki nie zostaną odkryte lepsze szkielety Australopithecus, Paranthropus i wczesnego Homo, klasyfikacje wielu skamieniałych homininów wokół tego kluczowego okresu ewolucji człowieka, około 2 milionów lat temu, najlepiej uznać za wstępne. W końcu Homo to tylko nazwa. Wiemy, że w pewnym momencie Homo wyewoluował z wcześniejszych australopitów. Im więcej znajdujemy skamieniałości, tym bardziej rozmyty jest podział na "małpoludów" (australopity) i "człowieka" (Homo). Skamieniałe homininy wyraźnie zlokalizowane we wczesnej części rodzaju Homo również przechodzą kryzys tożsamości. H. habilis została założona w 1964 roku przez Louisa Leakeya, Philipa Tobiasa i Johna Napiera. Wyznaczyli nowy gatunek na podstawie kości rąk i fragmentów czaszki młodocianego (OH 7), które wydobyli z Bed I w wąwozie Olduvai. H. habilis dosłownie oznacza "złota rączka" - imię, które opiera się na wywnioskowanych możliwościach chwytu skamieniałej ręki na podobieństwo człowieka. Gatunek został uznany za odpowiedzialny za narzędzia z wczesnej epoki kamienia (przemysł starodawny) znalezione w pobliżu stanowiska. H. habilis to gatunek homininów o małych mózgach, małych ciałach, które zwykle są trzymane oddzielnie od innych gatunków, które obejmowały ten sam zakres czasowy (2,3-1,8 mln lat temu), ale miały większy mózg, większe zęby i bardziej płaską twarz, H. Rudolfensis. Dwie skamieniałości symbolizujące różnice między H. habilis i H. rudolfensis to bardzo mała czaszka KNM-ER 1813 (500 cm3) i znacznie większa czaszka KNM-ER 1470 (775 cm3). Dla niektórych paleoantropologów (splitterów) różnica wielkości między nimi jest zbyt duża, aby utrzymać je w jednym gatunku, ale inni (ludzie) uważają, że jest to normalna zmienność godna jednego zmiennego gatunku. W 2006 roku nowe daty dla osadów w Koobi Fora poinformowali Patrick Gathogo i Frank Brown. Teraz mniejsza czaszka (1813) jest uważana za 0,25 Mya młodszą (1,65 Mya) niż większa (1,9 Mya). Jeśli te daty zostaną potwierdzone, problem może już nie występować.
Początki Człowieków (33)
PARANTROPUS (SOLIDNE AUSTRALOPITY)
Coraz popularniejsze staje się odwoływanie się do krzepkich gatunków australopitów z ich własnym rodzajem Paranthropus. Zgodnie z zasadami klasyfikacji Linneusza grupy muszą mieć jednego przodka, ale możliwe jest, że silne australopity wyewoluowały we wschodniej i południowej Afryce niezależnie od australopitów w każdym regionie (odpowiednio A. afarensis i A. africanus), co oznacza, że mogły mieć oddzielne korzenie. Ale dla jasności i ciągłości z obecnymi trendami, silne australopity są tutaj określane jako Paranthropus. Chociaż morfologiczne powiązania między australopitami a Paranthropusem są oczywiste, nie można pomylić Paranthropusa z ich odrębną morfologią czaszki i zębów. Paranthropus, który obejmuje Paranthropus aethiopicus, Paranthropus boisei i Paranthropus robustus, stanowi ewolucyjny ślepy zaułek w filogenezie homininów. Jest to wymarły rodowód, który rozgałęził się od australopitów około 2,5 miliona lat temu, a następnie zawisł na tyle długo, by koegzystować z wczesnym Homo we wschodniej i południowej Afryce do około 1 miliona lat temu. Pojemność czaszki Paranthropus waha się od 410 do 530 cm3. Chociaż dzieliły one powiększony rozmiar mózgu i dwunożność z A. africanus i A. afarensis, gatunki Paranthropus miały znacznie większe zęby i szczęki. Paranthropus używał swoich dużych płaskich zębów trzonowych w dużych szczękach do przetwarzania pokarmów włóknistych. Cztery zęby trzonowe współczesnego człowieka pasowałyby do jednego zęba trzonowego Paranthropusa, ale Paranthropus był tylko o połowę niższy niż przeciętny dorosły mężczyzna. Ich czaszki mogą pochwalić się wyraźnymi przyczepami do mięśni żucia (temporalis), w tym strzałkowym grzebieniem wzdłuż górnej części czaszki (podobnie jak mały kostny irokez). Ich duże, rozszerzone kości policzkowe i zwężenie oczodołowe robią miejsce, aby pomieścić te same mięśnie biegnące od szczęki do strzałkowego grzebienia, które byłyby wielkości funta steku po obu stronach głowy. Na podstawie morfologii czaszki i zębów, a także analizy izotopowej skład szkliwa zębów, wiemy, że Paranthropus żywił się twardymi, włóknistymi pokarmami i twardymi nasionami, orzechami i owocami. Średnia masa ciała dorosłego osobnika wynosiła około 33 kg dla kobiet i 45 kg dla mężczyzn przy wzroście porównywalnym z A. afarensis. P. boisei (wymawiane jako boy-zee-eye) jest prawdopodobnie najbardziej znanym przedstawicielem tego rodzaju, ponieważ może pochwalić się kompletną czaszką (OH 5), którą Louis Leakey znalazł w Olduvai Gorge w 1959 roku. (OH oznacza "Olduvai Hominid"). Leakey pierwotnie umieścił OH 5 w nowym rodzaju Zinjanthropus i zyskał przydomek "Zinj". Czaszka OH 5 była pierwszym homininem, który został dokładnie datowany (1,75 miliona lat temu) metodą potas/argon. Spośród trzech gatunków Paranthropus, P. boisei ma prawdopodobnie najbardziej przesadzone zęby trzonowe w porównaniu do wielkości małych siekaczy i ma "hipersolidny" grzebień czaszkowy dla mięśni żucia, co dało przydomek "człowiek dziadek do orzechów". Gatunki wahały się od 2,5 do 1 miliona lat temu, a okazy P. boisei występują również w Tanzanii i Koobi Fora w Kenii. Skamieniałości P. robustus pochodzą z południowoafrykańskich stanowisk Sterkfontein, Swartkrans i Drimolen, a ich czaszki i zęby przypominają mniej przesadzone wersje P. boisei. Obejmowały one od około 2,0-1,5 mln lat temu. P. robustus jest powiązany z zapisem narzędzi kostnych w miejscu jaskini Swartkrans. W pierwszej połowie XX wieku Raymond Dart zinterpretował te narzędzia jako broń, która była częścią kultury "osteodontokeratycznej" (z rogiem kostno-zębnym) używanej przez "zabójczych" małpoludów. Jednak od czasów Darta poczyniono znacznie więcej obserwacji zachowań szympansów i stały się dostępne nowe techniki mikroobrazowania artefaktów i kości. Przy współczesnej wiedzy jest jasne, że kościane narzędzia ze Swartkrans były używane do wykopywania bulw z ziemi, a niektóre były również używane jako różdżki do wyławiania termitów z ich kopców, podobnie jak szympansy pozyskują owady za pomocą gałązki. Pięknie kompletna czaszka, którą Alan Walker odkrył na początku lat 80. po zachodniej stronie jeziora Turkana w Kenii, jest najlepiej znanym okazem najwcześniejszego gatunku, P. aethiopicus (2,7-2,3 mln lat temu). Znany jest jako "Czarna Czaszka" (KNM-WT 17000) ze względu na kolor, który stał się skamieniały. (KNM oznacza Kenijskie Muzeum Narodowe, a WT oznacza West Turkana.) Czaszka przypomina aerodynamiczny, przylegający do drogi samochód sportowy ze względu na swoją prognatyczną twarz (jak maska) i ostre grzebienie czaszki, szczególnie poziome z tyłu głowa (jak spoiler). Szacuje się, że rozmiar mózgu P. aethiopicus był bardzo mały, około 410 cm3 i jest uważany za przodka P. boisei i P. robustus. Nie ma jasnego wyjaśnienia, dlaczego Paranthropus wyginął około 1 miliona lat temu. Intrygujące jest to, że grupa małych dwunożnych wegetarian wyginie, gdy wyższy, mięsożerny H. erectus rozprzestrzenił się w Starym Świecie. Cóż za uderzająca paralela do dzisiejszego znikania małp człekokształtnych w obecności ludzi.
Początki Człowieków (32)
Australopithecus anamensis
Najstarszym australopitem jest A. anamensis. Skamieliny A. anamensis pochodzą z okresu od 4,2 do 3,8 mln lat temu i pochodzą z Kanapoi i Allia Bay w Kenii oraz Asa Issie w Etiopii. Dziś kenijskie stanowiska znajdują się w pobliżu brzegu południowego krańca dużego jeziora Turkana. W różnych okresach w przeszłości części jeziora były znacznie zmniejszane, a miejsca te znajdowały się w dawnych deltach i obszarach zalewowych stale zmieniającego się krajobrazu. Kiedy w połowie lat 90. Meave Leakey, Kamoya Kimeu i Alan Walker odkryli pierwsze skamieliny A. anamensis, były to najwcześniejsze znane skamieniałości dwunożnych homininów. Żuchwa z Allia Bay ma kształt litery U małpy. Ma dużą kostną podporę na spojeniu żuchwy (tj. połączenie dwóch połówek żuchwy tuż pod siekaczami), aby chronić ją przed złamaniem jak kość wahadłowa podczas żucia. Ma większe zęby niż szympansy, a później A. afarensis. A. anamensis wykazuje dymorfizm płciowy podobny do goryla w wielkości kła i wielkości ciała. Istnieje jedna fragmentaryczna kość ramienna z Kanapoi, której brakuje morfologii chodzenia po kostkach małp człekokształtnych afrykańskich. Promień jest dłuższy niż nawet najdłuższy promień szympansa. Piszczel jest zorientowany w kostce jak współcześni ludzie, co wyraźnie wskazuje na użycie lokomocji dwunożnej. Dalsze dowody na to, że skamieliny A. anamensis nie należą do linii szympansów, pochodzą z czaszki w pobliżu ucha, gdzie otwór na strunę bębenkową (nerw twarzowy dla smaku) przechodzi przez kość inaczej niż u szympansów. Zęby i kości A. anamensis są bardzo podobne, ale wyglądają prymitywnie w porównaniu z późniejszym A. afarensis. Dlatego A. anamensis jest powszechnie uważany za przodka A. afarensis.
Australopithecus afarensis
Australopit o najbogatszym zapisie kopalnym to A. afarensis, a najbardziej znanym okazem jest częściowy szkielet pieszczotliwie nazywany "Lucy" (AL 288-1). Została odkryta przez Donalda Johansona w 1974 roku na stanowisku Hadar w regionie Afar w Etiopii i datuje się na około 3,2 miliona lat temu. Szacuje się, że 40 procent jej szkieletu jest zachowane, jeśli nie uwzględniono brakujących kości dłoni i stóp. W innym spektakularnym znalezisku w Hadar odkryto kilka osobników, które zmarły razem 3,2 miliona lat temu, a ta grupa A. afarensis nazywana jest Pierwszą Rodziną, ponieważ potwierdziło, że gatunek żył w grupach społecznych, ale przyczyna śmierci jest wciąż niejasna. A. afarensis żył od około 4 do 2,5 mln lat temu na obszarze rozciągającym się od Etiopii przez Tanzanię do Czadu. Skamieniałości A. afarensis znajdują się na stanowiskach, które niegdyś były głównie otwartymi i suchymi sawannami i lasami. Pojemność czaszki waha się od 400 do 500 cm3, a masa ciała dorosłych wynosi średnio 29 kg dla kobiet i 45 kg dla mężczyzn. Samce były prawie dwa razy większe od samic, podobnie jak współczesne goryle. Mieli długie ręce, krótkie kciuki i zakrzywione palce u rąk i nóg. Zęby mają pewne cechy przypominające małpy, w tym duże spiczaste kły i małą diastemę w dolnej szczęce, aby pomieścić górne kły. Wzory guzków trzonowych mają charakter pośredni między małpami a ludźmi, a same guzki są niskie i okrągłe jak u ludzi. Ogólnie zęby są mniejsze niż u małp człekokształtnych, a rząd zębów nie jest tak równoległy jak u małp człekokształtnych lub A. anamensis. Część pozaczaszkowa A. afarensis charakteryzuje się cechami podobnymi do małp, takimi jak krótkie nogi, długie ramiona i klatka piersiowa w kształcie lejka. Ale miał również ludzkie przystosowania do chodzenia w pozycji wyprostowanej, jak miednica w kształcie miski i mniej skrzywionych palców u rąk i nóg niż małpy (choć nadal zakrzywione). Ta mieszanka cech przypominających małpy i ludzi jest tym, co sprawia, że A. afarensis jest klasycznym łącznikiem między małpami a człowiekiem - prawdziwą dwunożną małpą. Jednak nadal toczy się debata na temat tego, jak nadrzewny pozostał ten gatunek. Dowody na dwunożność są znane z dużo większej liczby regionów anatomicznych niż u poprzednich gatunków, częściowo ze względu na lepszą ochronę i czystą liczbę odnotowaną, ale także dlatego, że dwunożność jest prawdopodobnie bardziej rozwinięta u A. afarensis. Jest dobrze zachowany staw kolanowy (koniec kości udowej i dopasowany szczyt kości piszczelowej) z Hadar, tego samego miejsca, w którym urodziła się Lucy, co wyraźnie pokazuje skośne kolano człowieka dwunożnego. Ślady przypisywane A. afarensis również wskazują na zaawansowany dwunożność. W miejscu Laetoli w Tanzanii, w twardej warstwie popiołu zachował się ślad homininów, datowany na około 3,7 mln lat temu, w czasie i zasięgu geograficznym A. afarensis. Mary Leakey prowadziła wykopaliska śladów wielu zwierząt, w tym królików i słoni. Te homininy, które przecinają drogę trójpalczastego konia, przypominają współczesnego człowieka. Występuje wgłębienie umożliwiające mocne uderzenie piętą, podobnie jak w nowoczesnym kroku dwunożnym, a także wygląd łuku, coś wyjątkowego dla ludzkiej stopy, które magazynuje energię jak sprężyna. Jednak duży palec jest nieco oddzielony od reszty palców.
Australopithecus africanus
Nazwa tego hominina dosłownie oznacza "południową małpę Afryki" i można ją znaleźć w jaskiniach, takich jak Swartkrans, Sterkfontein i Makapansgat w Afryce Południowej. A. africanus pojawił się po raz pierwszy około 3 milionów lat temu i żył do około 2,5 miliona lat temu w rejonach otwartych lasów i łąk. Te homininy są podobne do A. afarensis pod względem wielkości i kształtu ich szkieletu oraz różnicy w wielkości ciała między samcami i samicami. Jednak czaszki mają zaokrąglone szczęki, mniejsze kły, bardziej płaskie twarze (mniej prognatyzmu) i nie mają grzebienia kostnego widocznego u A. afarensis czy Paranthropus. Ich twarze mają charakterystyczne filary po obu stronach nosa. Ponadto ich mózgi były większe - średnio około 500 cm3. Ze względu na podobieństwo do późniejszych homininów, A. africanus jest często uważany za przodka wczesnego Homo. Najbardziej znaną skamieniałością A. africanus jest dziecko z Taung, czaszka i częściowy odlew wewnętrzny. Odkryta w Południowej Afryce w 1924 roku, ta skamielina została początkowo odrzucona jako ludzki przodek, ponieważ społeczność naukowa w tamtym czasie zakładała, że duże mózgi były pierwszą cechą, która ewoluowała u ludzi. Pomimo małego mózgu zęby dziecka Taung są podobne do ludzkich. Dodatkowo, lokalizacja otworu magnum wskazuje, że czaszka znajdowała się bezpośrednio na ciele i sugeruje, że osoba chodziła wyprostowana. Autentyczność znaleziska Taunga nie była powszechnie akceptowana aż do lat 40. XX wieku, kiedy odkryto dorosłe skamieniałości o tej samej mieszance cech. Dalsza inspekcja Dziecka z Taung pokazuje interesujące cechy, które są spójne z innymi australopitami. Kość czołowa (czoło) cofa się mniej niż u małpy, kły liściaste ("mleczne" lub "dziecięce") są mniejsze jak człowiek. Na podstawie harmonogramu wyrzynania się zębów szacuje się, że ma ono około trzech lub czterech lat. Rozmiar ciała, gdyby był ekstrapolowany na dorosłego, byłby jak szympans. Zmierzono objętość odlewu wewnętrznego na około 400 cm3, co w przypadku dorosłości do rozmiaru dorosłego wynosi około 450 cm3. Szkielet pozaczaszkowy A. africanus wykazuje wyraźne przystosowania do dwunożności. Sts 14, znaleziony w 1947 roku, to częściowy szkielet z kręgosłupem, miednicą i częścią kości udowej ze Sterkfontein, datowany na około 2,5 miliona lat temu. Kręgi wykazują wyraźne przystosowanie do dwunożności i mają ludzką miednicę w kształcie misy. Niedawno Ron Clarke odkrył zdumiewająco kompletny szkielet w jaskini Sterkfontein w Afryce Południowej, datowany na czasy A. africanus. Osobnik ten stał się znany jako "Mała Stopa" (STW 573), ponieważ jego stopę odkryto wcześnie, zanim reszta szkieletu została wykopana ze skały. Jest jednak jasne, że Littlefoot zostanie ostatecznie ujawniony jako najbardziej kompletny okaz wczesnego hominina w historii.
Australopithecus garhi
Nazwany "niespodzianką" w lokalnym języku afarskim ze względu na intrygującą kombinację cech, A. garhi jest znany z jednego stanowiska w Etiopii o nazwie Bouri w Środkowym Awash. Datuje się na 2,5 miliona lat temu i został ogłoszony przez Berhane′a Asfawa, Gen Suwę i Tima White′a w 1999 roku. Kości A. garhi są znaczącym odkryciem, ponieważ w tym okresie w zapisie kopalnym homininów z Afryki Wschodniej jest bardzo niewiele niesolidnych australopitów, a także jest to mniej więcej czas, w którym Homo dopiero zaczynał się tam rozwijać. A. garhi jest znany z czaszki i niektórych kości pozaczaszkowych. Czaszka ma strzałkowy grzebień i pojemność czaszki 450 cm3. Szczęka jest zaskakująco duża i solidna. Ma prymitywną morfologię czołową, twarzową i podnosową (poniżej nosa) w stosunku do A. africanus i wczesnego Homo. Zęby są jak gigantyczne wczesne zęby Homo. Siekacze są bardziej zbliżone rozmiarem do zębów trzonowych w porównaniu do wcześniejszych australopitów - stan, który jest również bardziej podobny do człowieka. Kości kończyn wskazują, że A. garhi miał niski wzrost jak wcześniejsze australopity. Były jednak bardziej umięśnione, ponieważ miejsca przyczepu mięśni na kościach są większe. Proporcje kończyn wskazują, że A. garhi miał stosunkowo długie przedramiona, jak wcześniejsze australopity, jednak kość udowa była znacznie dłuższa, co jest adaptacją dwunożną podzielaną przez Homo. Szczątki Bouriego dokumentują najwcześniej znany wzrost długości kości udowej, co wskazuje na wyraźną selekcję dla bardziej wydajnej dwunożności. Proporcje są mozaiką morfologii małpy i człowieka, ponieważ stosunek kości ramiennej do promienia/łokciowej jest podobny do małpy, ale stosunek kości ramiennej do kości udowej jest podobny do ludzkiego. Szczątki fauny z Bouri mają ślady po nacięciu kamiennych narzędzi, ale z tego miejsca nie ma żadnych narzędzi kamiennych. A. garhi jest nie tylko pierwszym homininem, który ma dłuższe nogi, ale także pierwszym, który kojarzy się z użyciem narzędzi do pozyskiwania wysoce pożywnego szpiku z kości zwierzęcych.
Kenyanthropus Platyops
Podobnie jak odkrycie Sahelanthropusa, nowe czaszki są czasami podstawą do chrztu nowego taksonu homininów. W przypadku Kenyanthropus platyops nowy rodzaj został nazwany od zniekształconej i silnie rozdrobnionej czaszki (KNMWT 40000). Meave Leakey kierowała zespołem naukowców, którzy zinterpretowali 3,5-milionową czaszkę z okolic jeziora Turkana, aby zaznaczyć początek opartego na diecie promieniowania adaptacyjnego - drugiej linii, która zaczyna się w środkowym pliocenie i obejmuje wczesny plejstoceński okaz Homo rudolfensis (KNM-ER 1470; ryc. 3.11, po prawej), z którym dzieli swoją charakterystycznie płaską twarz. Ta płaska twarz, a co za tym idzie znaczenie nazwy "platyops", wiąże się z inną dietą niż A. afarensis - jedyny inny hominin obecny w Afryce Wschodniej w wieku 3,5 miliona lat temu. Krytycy sprzeciwiają się wykorzystywaniu pojedynczej, mocno popękanej i rozdrobnionej czaszki K. platyops do ustanowienia całkowicie nowej linii homininów w czasach australopitów. Obecnie takson ten znajduje się w swoistym paleoantropologicznym okresie próbnym, czekając na poparcie z przyszłych znalezisk i analiz skamieniałości.
Początki Człowieków (31)
AUSTRALOPITEKI
Przedstawiciele rodzaju Australopithecus (często określani jako "australopiteki" lub "australopity") są niewątpliwie dwunożne, a niektóre gatunki są uważane za bezpośrednich przodków człowieka. Podobnie jak najwcześniejsze homininy, australopity i ich potomkowie Paranthropus występują tylko w Afryce. Dzięki ogromnym zapisom kopalnym australopiteków, z tysiącami okazów, w tym prawie kompletnymi czaszkami i szkieletami, wiele wiadomo o rodzaju. Wszystkie australopity mają wspólne cechy szkieletu. Mają mniejsze mózgi niż współcześni ludzie, mniej więcej o połowę mniejsze niż szympansy i goryle. Dorośli mieli około 130 cm wzrostu. Przypominały wyprostowane szympansy i mogły, ale nie musiały, wytwarzać kamienne narzędzia, ale podobnie jak żywe szympansy i goryle prawdopodobnie często używały narzędzi. Australopity znane są głównie ze skamieniałości czaszki i zębów. Kości czaszki za oczami wykazują bardzo silne zwężenie ("zwężenie zaoczodołowe"), co daje miejsce na duże mięśnie żucia (mięśnie skroniowe), które przyczepiają się od szczęki do czubka głowy. Ludzie mają bardzo małe mięśnie żucia i nie mają zwężenia oczodołu. Twarze australopitów są prognatyczne, a największa szerokość czaszki znajduje się u podstawy, w kierunku szyi, w przeciwieństwie do ludzi, których okrągłe czaszki są najszersze w górę. Na ogół australopity mają małe siekacze i kły w stosunku do masy ciała w porównaniu z żywymi małpami. W ten sposób są bardziej podobni do ludzi, ale nadal mają większe zęby w porównaniu do ludzi. Mają grubo emaliowane zęby trzonowe z bulwiastymi guzkami zakotwiczonymi w grubych, kostnych żuchwach. Rozmiar ciała australopiteka mieści się w zakresie współczesnych szympansów o wysokim stopniu dymorfizmu płciowego. Mieli podobne do ludzkich zdolności chwytu widoczne w ich kościach dłoni. Dyskutuje się, czy australopiteki były nadal biegłymi we wspinaczce po drzewach, czy też były obowiązkowymi dwunożnymi, ponieważ zachowały pewne cechy nadrzewne, takie jak klatka piersiowa w kształcie lejka, długie zakrzywione paliczki dłoni i stóp, stosunkowo krótkie nogi, stosunkowo długie ramiona. Ale mają też dowody na dwunożność, takie jak wyraźny wzór uderzania stopą (co widać po śladach Laetoli w Tanzanii), szerokie, rozszerzone kości biodrowe, kąt noszenia kości udowej, otwór magnum schowany pod czaszką, mniej przypominający kciuk palec, który jest bardziej zgodny z resztą palców. Istnieją cztery gatunki australopiteków, które są ogólnie akceptowane przez większość badaczy. Rodzaj Australopithecus powstał w pliocenie już 4,2 miliona lat temu z Australopithecus anamensis i trwa do około 2,5 miliona lat z Australopithecus garhi, gatunkiem, który pokrywa się z pochodzeniem naszego rodzaju Homo. Trzy gatunki australopitów występują w Afryce Wschodniej, a jeden, A. africanus, występuje w jaskiniach w Afryce Południowej. Należy zauważyć, że jaskinie w Afryce Południowej, w których znajdują się australopiteki, Paranthropus i niektóre okazy wczesnego Homo, nie zawierają żadnych dowodów na to, że homininy zamieszkiwały te jaskinie. Zamiast tego wydaje się, że ich kości gromadziły się, podobnie jak kości innych zwierząt, spadając przez szczeliny i szczeliny w ziemi do podziemnych jaskiń wapiennych. O wiele z tych nagromadzeń kości obwinia się drapieżniki, takie jak lamparty i duże ptaki drapieżne, ponieważ mają tendencję do używania drzew rosnących w pobliżu otworów. Są to te same rodzaje jaskiń, utworzone przez wody gruntowe rozpuszczające podłoże skalne, które są powszechne w Ameryce i które mogą ostatecznie tworzyć zapadlisko. Chociaż zaobserwowano naczelne inne niż ludzie, takie jak pawiany, szukające schronienia w jaskiniach, prawdziwe dowody na zamieszkiwanie przez ludzi jaskini nie pojawiają się aż do środkowego plejstocenu z archaicznymi ludźmi.
Początki Człowieków (30)
NAJWCZEŚNIEJSZE HOMININY
Do niedawna niewiele istotnych dowodów kopalnych pochodziło z późnego miocenu, kiedy to rozeszło się pochodzenie szympansów i ludzi. Teraz jest kilku pretendentów do tytułu najwcześniejszego hominina - Sahelantropus, Orrorin i Ardipithecus - i będzie potrzeba więcej odkryć skamieniałości dla paleoantropologów, aby zrekonstruować korzeń drzewa hominina. Niektóre pytania, jakie paleoantropolodzy zadają na temat najwcześniejszych homininów, to: Czy były to stworzenia leśne? Jeśli tak, to czy dwunożność rozwinęła się w lesie, w przeciwieństwie do jałowej sawanny, gdzie, jak się zakłada, ewoluował? Ile było gatunków wczesnych homininów? Czy pierwsze oddzielenie od LCA nastąpiło w wyniku specjacji sympatrycznej czy allopatrycznej?
Sahelanthropus
Francuski paleoantropolog Michel Brunet jest dobrze znany z tego, że krzyczy do mediów: "Wiedziałem, że to znajdę!" po odkryciu skamieniałości nazwał Sahelanthropus tchadensis. Jest to pojedyncza czaszka (czaszka bez żuchwy) datowana na 7-6 milionów lat temu. Znaleziony w suchym basenie jeziora Czad w pobliżu Toros-Menalla, S. tchadensis oznacza "małpę Sahel z Czadu". Skamielina została nazwana "Toumai", co w lokalnym języku oznacza "nadzieję na życie". Dzięki zegarom molekularnym, które oszacowały, że LCA żyła między 8 a 4 milionami lat, paleoantropolodzy tacy jak Brunet, którzy poszukują najwcześniejszych homininów, są bardziej pewni badania osadów starszych niż 5 milionów lat temu. Powiązane szczątki fauny wskazują, że region był kiedyś lasem, zupełnie innym niż piaszczysta pustynia, którą jest dzisiaj. Ponieważ nie ma szkieletu pozaczaszkowego, anatomia nie wykazuje bezpośrednich dowodów na dwunożność. Ma jednak inne cechy z późniejszymi homininami. Czaszka ma stosunkowo małe zęby, mało prognatyzmu i ma otwór magnum pod czaszką. Nie ma też podobnego do małpy kompleksu honowania kłów (CP3), który jest diastemą (lub przerwą) między kłem a przedtrzonowcem żuchwy, aby pomieścić i wyostrzyć duży kieł górnej szczęki, gdy szczęki są zamknięte razem. Jednak Sahelanthropus ma małpopodobne zęby, a także wiele małpopodobnych aspektów czaszki. Pojemność czaszki przy 350 cm3 jest niewielka, ale niekoniecznie różni się od tego, czego można by się spodziewać u najwcześniejszego hominina, ani od wczesnego przodka szympansa czy goryli. Odkrywcy wraz z wieloma innymi naukowcami uważają Sahelantropa za wczesnego hominina, co czyni go najwcześniejszym okazem w linii ludzkiej. Ale istnieje krytyka, że ponieważ nie ma szkieletu pasującego do czaszki, nie wiadomo jeszcze, czy ten gatunek był dwunożny. Co więcej, mosty nadoczodołowe są znacznie większe niż oczekiwano. Pierścienie nadoczodołowe nie stają się duże aż do 2 milionów lat temu z H. erectus, a wcześniejsze homininy nie mają dużych brodawek nadoczodołowych. Jednak goryle i szympansy mają grzbiety nadoczodołowe, a niektórzy krytycy twierdzą, że Sahelanthropus jest bardziej skamieniałą małpą. Datowanie Sahelanthropusa jest również trudne do interpretacji, ponieważ w morzu piasku, jakim jest Sahara, nie ma osadów wulkanicznych. Istnieje tylko jeden gatunek świni, który koreluje z datowanym miejscem Lothogam w Kenii, a korelacja biostratygraficzna wskazuje na wiek bliższy 6 milionów lat, a nie 6,5 lub 7 milionów lat temu. Jeśli Sahelanthropus rzeczywiście ma 6 milionów lat to jest bardzo podobny wiekiem do wschodnioafrykańskiego hominina Orrorin tugenensis. Niestety niewiele zachowanych części pokrywa się w celu bezpośredniego porównania, więc potrzeba więcej skamielin, aby lepiej zrozumieć związek między tymi najwcześniejszymi homininami.
Orrorin
Najwcześniejsze postczaszkowe skamieliny homininów pochodzą z 6 milionów lat temu. Obejmują one kilka fragmentów kości nóg i kilka zębów Orrorin tugenensis ze wzgórz Tugen w regionie jeziora Baringo w środkowej Kenii. Brigitte Senut i Martin Pickford dokonali odkrycia na przełomie XIX i XX wieku, co zainspirowało jego przydomek "Człowiek milenijny". Orrorin jest podobny do człowieka pod względem grubości szkliwa zębów, wielkości zęba i morfologii. Stwierdzono również, że jest dwunożny ze względu na morfologię kości udowej. Szyja kości udowej jest długa jak u ludzi. Ponadto zdjęcia rentgenowskie wskazują, że infrastruktura kostna bardziej przypomina ludzi niż małpy. Oznacza to, że kość została zbudowana, aby wytrzymać rodzaje sił generowanych podczas chodzenia w pozycji wyprostowanej na dwóch nogach. Dwunożny status Orrorinu jest jednak wciąż przedmiotem dyskusji, ponieważ szczątki skamieniałości nie dają jeszcze jednoznacznego, przekonującego dowodu.
Ardipithecus
Pierwotnie umieszczony w Australopithecus, Ardipithecus to rodzaj reprezentowany przez gromadzącą się liczbę okazów z miejsc takich jak Aramis w regionie Middle Awash w Etiopii. Ardipithecus ramidus oznacza "małpę korzeniową" zarówno po łacinie, jak i w ojczystym języku regionu. Zmiany morfologiczne od 5,8 do 4,4 milionów lat temu są niewielkie, więc ci, którzy pracują nad tymi skamieniałościami (np. Tim White, Gen Suwa, Berhane Asfaw, Johannes Haile-Selassie) zachowali zmienność na poziomie podgatunku z Ardipithecus ramidus ramidus (4,4 miliona) i Ardipithecus ramidus kadabba (5,8-5,2 mln lat). Niewiele zachowała się morfologii czaszki i części pozaczaszkowej Ardipithecus. Na opublikowanie czeka fragmentaryczny szkielet, ale na razie w aktach są tylko fragmenty czaszki, zębów, szczęk i kilka szczątków pozaczaszkowych. Zmniejszone rozmiary górnych i dolnych kłów w stosunku do przedtrzonowców i trzonowców są bardziej ludzkie niż małpie. Jednak Ardipithecus ma prymitywnie cienką emalię i prymitywną, choć zmniejszoną morfologię kompleksu honującego CP3. Czy Ardipithecus był dwunożny? Otwór magnum znajduje się pod czaszką w A.r. ramidus, co sugeruje, że tak. Istnieje również pojedyncza kość palucha (falanga) A.r. kadabba, która ma kształt człowieka, w przeciwieństwie do szympansa. Dowody z miejsc Ardipithecus - takie jak nasiona, małpy i antylopy wędrowne - wskazują, że siedliska były w większości lasami o zamkniętych baldachimach, a nie sawannami.
Początki Człowieków (29)
CZWORONOŻNY DO DWUNOŻNEGO
Wiele zmian zachodzących w linii homininów wiąże się z przystosowaniem się do dwunożnego ciała czworonożnej małpy. Zbudowane do wspinania się, huśtania się i przedzierania się przez drzewa, ciała małp mają krótkie, płaskie tułowia, szerokie biodra, długie obojczyki, łopatki z tyłu zamiast z boku ciała (jak małpa lub pies), okrągła głowa kości ramiennej, która zapewnia elastyczny mankiet rotatorów na ramieniu, półwyprostowaną postawę, długie ramiona, krótkie nogi, wydłużone kończyny przednie, długie zakrzywione palce i brak ogona. Ludzie nadal posiadają wiele cech przypominających przodków małp, które były trampoliną nie tylko do ewolucji dwunożności, ale także ewolucji rzucania przez ramię i wytwarzania narzędzi, co częściowo zależy od zachowania części tej rozgałęzionej anatomii. Główne zmiany w proporcjach kończyn nastąpiły z powodu dwunożności, ponieważ lepiej mieć długie nogi do chodu, ludzie wykształcili znacznie dłuższe nogi w stosunku do wielkości ciała niż małpy. Mamy też znacznie krótsze ręce niż
małpy w stosunku do naszego ciała, a kilka z powodów to prawdopodobnie większa kontrola podczas używania narzędzi i rzucania, a także optymalna długość ramion do kołysania ramion na bok podczas chodzenia i biegania. Najwcześniejsze homininy miały podobny plan budowy ciała do żywych goryli, szympansów i bonobo, i toczy się dyskusja, czy chodziły na knykciach tak jak one. Nadgarstek poruszający się po kostkach musi być mocny i sztywny, aby utrzymać ciężar zwierzęcia, więc morfologia kości promieniowej pokazuje mechanizm blokujący, aby zapobiec upadkowi nadgarstka. Mogą istnieć anatomiczne pozostałości adaptacji chodzenia po kostkach u wczesnych australopiteków, ale kości ramion i nadgarstków jest niewiele, a cechy wydają się być pośrednie. Dowody na wcześniejsze homininy, z mniejszymi adaptacjami dwunożnymi (i przypuszczalnie większą szansą na zachowanie takich adaptacji, gdyby były tam w pierwszej kolejności) nie zostały jeszcze odkryte. Parsimony wspiera LCA jako kastety. Jeśli wspólny przodek goryli i
szympansów był chodzącym po kostkach, to ma sens, że LCA ludzi i szympansów też był taki, i że ludzie stracili przystosowanie do chodzenia po kostkach, gdy wyewoluowali dwunożność. Natura nie zawsze jest posłuszna naszej zasadzie skąpstwa, więc możliwe jest również, że chodzenie po kostkach powstało niezależnie zarówno w linii rodowej goryli, jak i szympansów, co oznacza, że możliwe jest, że LCA nie wykazywała jeszcze zachowania chodzenia po kostkach. Analizy genetyczne wykazały, że szympansy i ludzie są bliżej spokrewnieni niż szympansy z gorylami, więc nie można jeszcze wykluczyć samodzielnego nabywania umiejętności chodzenia po kostkach u małp człekokształtnych. Więcej skamielin przodków małp człekokształtnych i ludzkich z przełomowego okresu późnego miocenu i wczesnego pliocenu ostatecznie rozwiąże kwestię chodzenia na kostkach homininów.
Początki Człowieków (28)
OSTATNI WSPÓLNY PRZODEK
Jak wyglądałby ostatni wspólny przodek? Jak wyglądałby najwcześniejszy hominin? Jak bardzo przypominałoby współczesnego szympansa? Paleoantropolodzy muszą wykazać, że ich skamieliny nie są skamieniałościami szympansów ani goryli, aby mogły zostać zaakceptowane jako część drzewa genealogicznego homininów. Jest to wysoki poziom dla wczesnej części zapisu kopalnego homininów (między 7 a 4 milionami lat temu), kiedy homininy były jeszcze bardzo podobne do małp. Poza tym nie ma skamieniałych szympansów ani goryli od późnego miocenu do pliocenu, które można by porównać. W zapisie jest tylko jeden rozpoznany kopalny szympans, datowany na stosunkowo niedawny środkowy plejstocen. Kilka zębów pobrano z miejsca w pobliżu jeziora Baringo w Kenii. Datowanie argonowo-argonowe ustaliło zęby na około 525 Kya. Wyglądają dokładnie jak zęby współczesnego szympansa i, co ciekawe, są tam skamieniali ludzie z tych samych regionalnych lokalizacji, którzy prawdopodobnie żyli w tym samym miejscu w pradziejach, co szympansy. Tropikalne siedliska szympansów i goryli są winne ich nieobecności w zapisie kopalnym. Współczesne małpy afrykańskie trzymają się ciepłych, wilgotnych lasów tropikalnych i zakłada się, że w przeszłości korzystały z tych samych siedlisk. Skamieniałość rzadko występuje gdziekolwiek, nie mówiąc już o tego rodzaju siedliskach, w których kwasowość gleby jest wysoka, a organizmy pożerające zwłoki są dokładne i wydajne. Trudno jest rozpoznać i właściwie zidentyfikować prymitywne gatunki w pobliżu głównych zdarzeń dywergencji ewolucyjnej, takich jak LCA i jego krewni. Co więcej, jak oceniamy zmienność wewnątrz i między gatunkami wymarłych stworzeń? Opieranie szacunków zmienności na tym, co dzisiaj widzimy u małp, małp człekokształtnych i ludzi, jest najbardziej logicznym i rozsądnym sposobem przezwyciężenia tego problemu, ale wiąże się również z ryzykiem zamaskowania różnych wzorców zmienności, które mogły istnieć w przeszłości. W paleontologii są inne przeszkody. Często skamieniałości są połamane lub rozdrobnione. Niektóre z najbardziej diagnostycznych anatomii czaszki są również najbardziej kruche. Aby złożyć fragmentaryczne kości, potrzebna jest wiedza z wieloletnich badań anatomicznych i doświadczenie, ale przy pomocy nowoczesnych technik obrazowania, takich jak skanowanie CT, proces ten staje się znacznie łatwiejszy. Dzięki wirtualnym rekonstrukcjom skamieniałości paleontolodzy są w stanie naprawić pęknięcia skamieniałości bez konieczności ich ręcznego czyszczenia, ekstrakcji i składania, co często jeszcze bardziej niszczy skamieliny. Liczba odkrytych skamieniałości młodocianych homininów może również prowadzić do problemów z ich identyfikacją. Osobniki, które nie są w pełni dorosłe lub w pełni dorosłe, nie mają wszystkich cech diagnostycznych ich gatunku. Duża część ważnych okazów homininów to niemowlęta lub dzieci, na przykład dziecko z Taung (Australopithecus africanus), chłopiec Nariokotome H. erectus, dziecko Dikika (Australopithecus afarensis), dziecko Mojokerto (Homo erectus), a jest ich kilka, młodocianych neandertalczyków. Ich wiek zależy od stadium wyrzynania się zębów. Zarówno zęby mleczne , jak i stały drugi zestaw zębów wyrzynają się według określonego wzoru i postępują zgodnie z określonym harmonogramem. Na przykład, wyrzynają się pierwsze zęby trzonowe człowieka w wieku około sześciu lat i tak dalej. Podobny wzór i harmonogram ma miejsce w przypadku łączenia płytek wzrostu na końcach kości długich. Różnice w wielkości ciała i wielkości przyczepów mięśniowych na kościach wskazują, czy dana osoba była mężczyzną czy kobietą, ale takie różnice są znacznie mniej wyraźne przed osiągnięciem dorosłości, więc określenie płci nieletnich jest trudne, im są młodsze. Ponieważ skamieniałości są reliktami budowy stworzeń, gatunki identyfikuje się na podstawie cech anatomicznych - głównie wysoce diagnostycznych cech czaszki i zębów. Ogólnie rzecz biorąc, istnieją trendy w zapisie kopalnym homininów, które możemy śledzić, aby śledzić podróż naszych przodków od LCA do nas.
Trendy na przestrzeni czasu w plio-plejstoceńskim zapisie kopalnym hominina
• Zmniejszenie rozmiaru twarzy
• Redukcja prognatyzmu, czyli projekcji twarzy
• Zmniejszenie wielkości mola, względne i bezwzględne
• Zmniejszenie wielkości kła
• Utrata cech nadrzewnych, takich jak długie zakrzywione palce u rąk i nóg, krótkie nogi i długie przedramiona
• Nabycie cech lądowych i dwunożnych, takich jak krótsze palce u nóg, dłuższe nogi (zwłaszcza kość udowa) i krótsze ramiona
• Zwiększenie pojemności czaszki (wykorzystywane do określenia rozmiaru mózgu)
• Zwiększenie okrągłości czaszki
• Zwiększ, a następnie zmniejsz w brwiach
• Wzrost wzrostu i wielkości ciała
• Zmniejszenie wytrzymałości kości
• Zmniejszenie dymorfizmu płciowego lub różnice między mężczyznami i kobietami w wielkości ciała oraz cechach zębów i czaszki
Początki Człowieków (27)
KRZEWY I DRZEWA
Dopóki skamieniałości nie otrzymają etykiet, które dokładnie opisują, do jakiego gatunku należały, co jadły, z kim się krzyżowały i czy mogły mówić, czy nie, zawsze będą spory o ich interpretację. Nie chodzi o to, czy skamieniałe homininy zawierają wskazówki dotyczące naszej ewolucji, ale o to, jak je zinterpretować i dojść do logicznego konsensusu co do tego, jak powstał Homo sapiens. Pierwszą przeszkodą w osiągnięciu takiego konsensusu jest to, że skamieniałości homininów są rzadkie. Jasne, trudno jest dziś odwiedzić jakąkolwiek część świata bez wpadnięcia na ludzi, ale nasza owadziopodobna kolonizacja Ziemi jest zjawiskiem bardzo niedawnym. Przed 10 Kya wszyscy ludzie żyli w małych grupach rzadko przekraczających 150 osobników. Tak więc szanse na znalezienie skamieniałości hominina są zmniejszone wyłącznie ze względu na wielkość populacji w przeszłości. Paleoantropolodzy znajdują znacznie więcej kości i zębów gryzoni i byków (antylop itp.) niż znajdują homininów, ale wciąż istnieją tysiące skamieniałości homininów. Z powodu dwóch fundamentalnie odmiennych poglądów na interpretację ewolucji istnieją argumenty, które skamieniałości należy włączyć do linii prowadzącej do człowieka, a które umieścić na bocznych gałęziach. Zasadniczo "rozszczepiacze" widzą filogenezę homininów jako krzak, ale "drzewcy" widzą ją jako drzewo. Najbardziej ekstremalni lumperzy widzą go jako kaktus saguaro, z może dwiema lub trzema liniami/oddziałami. Krzaczasty pogląd przedstawia bardzo skomplikowaną historię filogenetyczną homininów z wieloma gatunkami istniejącymi w dowolnym czasie. Drzewo lub kaktus symbolizuje pogląd, że homininy ewoluowały z bardzo małą różnorodnością i bardzo niewielkim nakładaniem się gatunków, w długich liniach ewolucyjnych. Rozłamowcy twierdzą, że zapis kopalny nie może zawierać przedstawicieli wszystkich gatunków homininów, jakie kiedykolwiek istniały; że musi istnieć tak duża różnorodność, z której nie pobiera się próbek ze względu na problemy związane z ochroną. Pogląd trębaczy jest konserwatywny, który wskazuje na dowody z zapisu kopalnego, jaki znamy. Argument krzak kontra drzewo jest ważny, jeśli mamy określić naturę ewolucji człowieka. Jest to również ważne dla tego, jak rekonstruujemy codzienne życie homininów 1,5 miliona lat temu. Życie może wyglądać zupełnie inaczej, gdyby istniał oddzielny gatunek dwunożnych o wysokości 3 stóp, dzielących z nami planetę, i tak wyglądało życie dla ludzkiego H. erectus, który współistniał z małpopodobnymi australopitekami i dla ludzi z Indonezji około 13 000 lat temu, kiedy na Flores mieszkali tak zwani "hobbici". A jeśli bycie jedyną dwunożną małpą używającą narzędzi, tak jak my teraz, jest raczej wyjątkiem niż regułą?
Dlaczego "Hominin"?
"Hominin" zastępuje wszechobecny niegdyś w literaturze termin "hominid". Oba terminy są używane w odniesieniu do ludzi i ich wymarłych przodków od czasu ostatniego wspólnego przodka z szympansami (LCA) i oba są nadal akceptowane. Różnica w terminach dotyczy tego, jak naukowcy postrzegają relacje między ludźmi a resztą małp człekokształtnych. Tradycyjnie ludzie byli jedynym gatunkiem należącym do rodziny Hominidae ("hominidów"), przy czym małpy człekokształtne umieszczano w osobnej rodzinie Pongidae, ponieważ podobieństwa anatomiczne grupują małpy człekokształtne z wyłączeniem ludzi. Jednak ze względu na podobieństwo genetyczne między szympansami, ludźmi i gorylami (z wyłączeniem orangutanów), szympansy, goryle i wszyscy ich skamieniali przodkowie są coraz częściej wrzucani do ludzi w Hominidae. Dlatego też stworzono podkategorie w celu dalszego zróżnicowania grup, w których ludzie i ich skamieniali przodkowie nazywani są homininami.
Początki Człowieków (26)
Skamieniałe Małpy Człekokształtne
Obecnie małpy znacznie przewyższają liczebnie małpy człekokształtne, ale w miocenie sytuacja uległa odwróceniu. W miocenie jest stosunkowo niewiele skamieniałości małp. W tamtym czasie małpy wypromieniowały się, wypełniając większość nisz, które małpy później przejęły i wypełniają dzisiaj. Różnorodność małp zmniejszyła się z biegiem czasu, tak że w pliocenie dominowały małpy, a na potwierdzenie tego istnieją bogate zapisy skamieniałości małp z całego Starego Świata. Możliwe, że małpy po prostu krzyżowały się z małpami. Wielkie małpy czekają ponad cztery lata między narodzinami, ponieważ ich potomstwo rozwija się znacznie wolniej niż małpy. Większość zapisów kopalnych małp człekokształtnych pochodzi z Afryki Wschodniej i Europy, a kilka gatunków znajduje się w Azji i Indonezji. Wczesne małpy człekokształtne wykazują wzrost rozmiarów ciała i różnorodność trybów lokomotorycznych w porównaniu z ich małpopodobnymi przodkami. Chociaż w zapisie jest wiele małp mioceńskich, żadna z nich nie jest z pewnością umieszczona w bezpośrednim rodowodzie prowadzącym do człowieka. Mozaikowa natura małp człekokształtnych, czyli ich mieszanka cech, która czyni je niepodobnymi do żadnych współczesnych małp człekokształtnych, stanowi wyzwanie dla paleontologów w ich interpretacji. W miocenie Afryka Wschodnia była pokryta lasami deszczowymi i wypełniona małpami o różnych rozmiarach i diecie. Te skamieniałe gatunki dzieliły trendy z żyjącymi małpami: duże mózgi, mniej kręgów, długie kończyny górne i brak ogonów. Ich czaszki i zęby pokazują, że jadły pokarmy włókniste, takie jak owoce, orzechy i twarda roślinność. Miały grube szkliwo zębów, zaokrąglone guzki, rozszerzone kości policzkowe dla mięśni żuchwy (skroniowych), aby zmieścić się pod spodem. Adaptacje pozaczaszkowe tych małp obejmowały anatomię zbudowaną z myślą o nadrzewności, wspinaczce i brachiacji. Tylko jedna małpa mioceńska ma uzgodniony przodek i potomek z żywą małpą i jest to Sivapithecus z 14 milionów lat temu na wzgórzach Siwalik w Pakistanie. Sivapithecus jest przodkiem współczesnych orangutanów. Jest wielu kandydatów, ale jak na razie nie ma zgody co do tego, który z nich jest przodkiem afrykańskich małp człekokształtnych: goryli, szympansów i ludzi. Najbardziej znanym gatunkiem małpy mioceńskiej jest Proconsul, znany głównie ze stanowisk na wyspie Rusinga w kenijskich wodach Jeziora Wiktorii, datowany na okres od 20 do 18 milionów lat temu. Istnieje ponad dwanaście częściowych szkieletów Prokonsula i kilka szczęk, izolowane kości i zęby oraz jedna dobrze zachowana czaszka. Najlepiej wyobrazić sobie Proconsula jako małpopodobnego ciała z małpimi cechami zębów (adaptacje owocożerne), czaszki, niektórych aspektów kończyn i pozbawionego ogona. Z pewnością brak dowodów na coś, zwłaszcza w kapryśnym zapisie kopalnym, nie dowodzi, że nie istnieje. Jednak żaden z częściowych szkieletów nie zachował kręgów ogonowych (ogonowych), a kość krzyżowa nie wydaje się mieć żadnych anatomicznych powiązań z ogonem. Ponieważ zakłada się, że ogony zostały utracone tylko raz w ewolucji człekokształtnych, brak ogona u Proconsula potwierdza jego status małpy. Liczne małpy mioceńskie o cechach małpoludów mogły być bezpośrednimi przodkami żyjących małp człekokształtnych. Morotopithecus z 21 Mya w Ugandzie jest interpretowany jako mający cechy rozgałęzione i zawieszone, podobnie jak małpy człekokształtne. W miarę zbliżania się skamieniałych małp człekokształtnych do środkowego miocenu, nabierają wielu bardziej nowoczesnych cech. Afrykańskie skamieniałe małpy Nacholapithecus, Kenyapithecus, Kamoyapithecus, Samburupithecus, Otavipithecus i Afropithecus są pretendentami do afrykańskich przodków małp człekokształtnych. Ale w Europie jest jeszcze więcej gatunków podobnych do małp. Około 12 Mya Dryopithecus pojawia się w Hiszpanii, Rumunii, Francji i na Węgrzech. Oreopithecus jest tak podobny do małpy, że interpretuje się go nawet jako przystosowany do początkowej dwunożności, ale te poglądy nie są mocno wspierane. Inne europejskie małpy człekokształtne to Ouranopithecus (Grecja), Ankarapithecus (Turcja), Griphopithecus i Grecopithecus. Najnowsze znalezisko, dobrze zachowany częściowy szkielet Pierolapithecus, ma wiele cech wspólnych z żyjącymi małpami człekokształtnymi. Rekord małpy mioceńskiej jest zagadkowy. Zakłada się, że skoro wszystkie małpy człekokształtne z wyjątkiem jednej żyją w Afryce, ewoluowały tam, ale skamieniałe małpy z Afryki są bardziej mozaiką cech przypominających małpy niż ich mniej małpopodobni, bardziej małpopodobni krewniacy w Europie . Po odkryciu większej liczby skamielin w późnym miocenie, zarówno w Afryce, jak iw Europie, historia ewolucji małp i pojawienia się linii homininów będzie znacznie jaśniejsza.
Gigantopithecus: prawdziwa wielka stopa
Być może legenda o Wielkiej Stopie ma taką siłę przetrwania ze względu na swoje starożytne korzenie. Plejstoceński zapis kopalny z Chin zawiera zęby i szczęki prehistorycznej Wielkiej Stopy zwanej Gigantopithecus blacki. Chociaż nie odkryto żadnych szkieletów (ani kości stóp), szacowany rozmiar ciała w oparciu o rozmiar zębów jest ogromny i wynosi ponad 400 kg, co jest ostrożnym szacunkiem. Nawet jeśli zęby były większe niż oczekiwano dla wielkości ciała, Gigantopithecus nadal był największym znanym naczelnym, jaki kiedykolwiek istniał. W przeciwieństwie do legendarnej Wielkiej Stopy, Gigantopithecus prawdopodobnie poruszałby się czworonożnie, jak inna podobna legendarna postać, King Kong. Olbrzymia małpa wymarła tylko około 100 Kya, więc miała wiele czasowych i geograficznych nakładań z Homo erectus. Goryle mają stopy podobne do ludzkich, ponieważ przystosowania do noszenia ich dużych, ciężkich ciał i spędzania czasu na ziemi faktycznie naśladują adaptacje ludzkich stóp, które przystosowały się do dwunożności i ziemskiego charakteru. W konsekwencji ślady goryli również wyglądają podobnie do ludzkich. Jeśli Gigantopithecus przypominał współczesne goryle, a prawdopodobnie nim był, jego stopy mogły być podobne do goryli, a może nawet do ludzi. Być może światowi tropiciele Wielkiej Stopy powinni udać się do Chin, aby znaleźć skamieniałe ślady naprawdę prawdziwego, choć starożytnego Bigfoota.
Początki Człowieków (25)
Najwcześniejsze naczelne i kopalne małpy
Zapis kopalny ssaków jest pełen fascynujących wymarłych zwierząt, których podzbiór składa się ze skamieniałości naczelnych. Najwcześniejsze naczelne występują w paleocenie i przypominają wiewiórki. Naczelne oddzieliły się od reszty ssaków bardzo wcześnie wraz z gryzoniami, więc skamieniałe naczelne są jednymi z najstarszych i najbardziej prymitywnych ssaków kopalnych w historii. Najwcześniejsze zwierzęta przypominające naczelne nazywane są plesiadapiformami (rodzina Plesiadapiformidae) i występują w skałach Europy i zachodniej Ameryki Północnej (szczególnie Wyoming). Skład skalny i związana z nim fauna kopalna wskazują, że plesiadapiformy preferowały ciepłe, wilgotne lasy, które w tym okresie dominowały w tym regionie. Podobnie jak większość wczesnych ssaków, plesiadapiformy były nocnymi czworonogami o dobrych zmysłach węchu - cecha ta wynika z dużych pysków i stosunkowo małych oczodołów, co wskazuje na stosunkowo mniejszą zależność od wzroku. Podobnie jak współczesne naczelne plesiadapiformy miały charakterystyczne zęby oraz chwytne dłonie i stopy. Ale w przeciwieństwie do współczesnych naczelnych miały pazury, brakowało im cech skaczących szkieletu (prymitywny dla naczelnych) i nie mieli dużych, zamkniętych, kostnych oczodołów. Okrytozalążkowe, czyli rośliny, które rozsiewają nasiona wraz z kwiatami i owocami, wyewoluowały w okresie kredowym, tworząc nowe źródło pożywienia dla zwierząt i niszę, z której korzystały najwcześniejsze naczelne. Były to zwierzęta nadrzewne, zjadające owoce, nasiona i owady. Trwa debata na temat tego, który produkt żywnościowy - ożywione owady (z których wiele zapylało rośliny okrytozalążkowe) czy kolorowe owoce - był siłą napędową ewolucji cech naczelnych, takich jak ostre ubarwienie i widzenie stereoskopowe oraz zręczne chwytanie rąk. Chociaż mogły to być zarówno owady, jak i owoce, ponieważ wszystkie współczesne naczelne jedzą różnorodne pokarmy i nie ograniczają się do jednej specjalizacji. W eocenie skamieniałości pierwszych w pełni rozpoznanych naczelnych, które wykazują cechy, których brakowało plesiadapiformom, żyły w Ameryce Północnej i Europie, które były wówczas połączone lądem. Omomyidae (z rodziny Omomyidae) były małymi owadożercami, przypominającymi współczesne wyraki. Adapidy (z rodziny Adapidae) były większymi owocożercami i liśćmi, przypominającymi współczesne lemury. Gdybyśmy byli lemurami, pominęlibyśmy poniższe sekcje dotyczące skamieniałych małp, małp człekokształtnych i ludzi, a podążalibyśmy skamieniałymi śladami naszych przodków od adapidów do naszego obecnego życia na Madagaskarze. Ale jesteśmy ludźmi, więc będziemy podążać skamieniałymi śladami naszych przodków, od najwcześniejszych małp naczelnych, przez małpy człekokształtne, aż wreszcie do ludzi (w tym znacznie większych, dwunożnych naczelnych, które również żyją na Madagaskarze). Wszystkie małpy, małpy człekokształtne i ludzie (haplorhine) mają wspólnego przodka z Azji we wczesnym eocenie, znanego jako Eosimius lub "małpa świtu". Eosimius jest niezwykle mały, wielkości kciuka człowieka, ale ma cechy, które odróżniają go od lorisów i lemurów i łączą go z małpami. Wiele z tego, co wiemy o wczesnej ewolucji haplorinów, pochodzi ze skamieniałości zachowanych na pustyni Fajum w Egipcie - jakieś 60 mil na południowy zachód od Kairu. Miejsca tam zostały po raz pierwszy wykopane na początku XX wieku i nadal są wykopywane przez zespoły kierowane przez Elwyna Simonsa. Te miejsca na skraju Sahary są dziś bardzo suche, ale są bogate w skamieniałe ssaki z czasów, gdy region ten był zalesionym bagnem 34 milionów lat temu. Prymitywne małpy Apidium i Aegyptopithecus pochodzą z Fajum. Apidium ma formułę dentystyczną platyrrhines (małpy Nowego Świata), ale Aegyptopithecus ma formułę katar (małpy Starego Świata, małpy człekokształtne i ludzie). Oba gatunki mają zrośnięte żuwaczki i całkowicie zamknięte kostne oczodoły. Zapis skamieniałości małp Starego Świata, które oddzieliły się od linii małp człekokształtnej i ludzkiej, jest bardzo bogaty w pliocenie i plejstocenie, ale wcześniej w ewolucji linii skamieniałości są rzadkie. Wyjaśnienie, w jaki sposób małpy dotarły do Ameryki Południowej, jest dużym wyzwaniem. Małpy Nowego Świata ewoluowały w oderwaniu od wspólnego przodka z małpami Starego Świata i od tego czasu stały się bardzo zróżnicowane. Małpy Fossil New World pojawiają się po raz pierwszy w Ameryce Południowej około 28 milionów lat temu. Niektóre z najwcześniejszych skamielin znalezionych w Boliwii są bardzo podobne do tych odkrytych w Fajum z oligocenu. Istnieją dwie możliwe drogi do Ameryki Południowej, ale obie wydają się trudne. Jedna droga prowadzi z Ameryki Północnej, ale zapis kopalny śledzący jakąkolwiek ewolucję małp w Ameryce Północnej w tym czasie jest bardzo nieliczny, a ponadto przesmyk w Panamie nie był powiązany z Ameryka Południowa jeszcze, więc musieliby przeskoczyć wyspę przez Karaiby. Druga możliwa droga prowadziłaby przez południowy Ocean Atlantycki z Afryki, gdzie znaleziono wczesne prekursory kopalne. Małpy Nowego Świata mogły unosić się na tratwach porośniętych zbitą roślinnością lub "pływających wyspach". Te dwa kontynenty były bliżej siebie, ponieważ dno morskie rozciągające się na grzbiecie środkowoatlantyckim było we wcześniejszej fazie odpychania Ameryk od Europy i Afryki (co nadal dzieje się z tą samą prędkością, z jaką rosną twoje paznokcie).
Początki Człowieków (24)
ZMIANY KLIMATU I PALEOŚRODOWISKO
Ewolucja homininów miała miejsce w czasach wielkich globalnych zmian klimatycznych. Odwiercone rdzenie głębinowe i lodowcowe zawierają historyczne zapisy dotyczące klimatu Ziemi. Każda warstwa lodu, która się utworzyła, i każda warstwa dna morskiego, która się osadzała, zawiera migawkę warunków klimatycznych podczas jego tworzenia. O tych warunkach mówi ilość określonych związków chemicznych w warstwach. Czasy chłodzenia (glacjały) lub ocieplenia (interglacjały) są wskazywane przez proporcje izotopów tlenu w zróżnicowany sposób przyswajane przez mikroskopijne organizmy jednokomórkowe (otwory lub "otwory"), które są skamieniałe w warstwach. Niektóre wzorce sugerują, że wymieranie, a nie specjalizacja, są bardziej skorelowane ze zmianą klimatu. Jednak wymieranie wynikające z takich zmian może otworzyć nisze do wypełnienia przez nowe organizmy. Po wyginięciu dinozaurów na granicy kredy i trzeciorzędu nastąpiło stopniowe powiększanie się ssaków, które ostatecznie wypełniły puste nisze dinozaurów o małych i dużych ciałach. Ewolucja najwcześniejszych naczelnych natychmiast wyprzedziła gwałtowne globalne ochłodzenie, które rozpoczęło się w eocenie. Inny długoterminowy trend chłodzenia i suszenia zbiegł się z pochodzeniem homininów między 8 a 5 milionów lat temu. Oscylacje w globalnym klimacie wystąpiły podczas ewolucji homininów i prawdopodobnie wpłynęły one na wynik ewolucji człowieka. Począwszy od późnego pliocenu, około 2,8 mln lat temu Afryka stała się chłodniejsza i bardziej sucha, co prawdopodobnie przekształciło lasy w łąki. Roślinność wschodnioafrykańska przeszła z zamkniętych zalesionych baldachimów do otwartych, suchych sawann o zmniejszonych i sezonowych opadach. Następnie, kontynuując dolny plejstocen (1,8 Mya-900 Kya), klimat światowy zaczął się szybciej ochładzać. Środkowy plejstocen (900-125 Kya) charakteryzował się zimnymi okresami zlodowacenia, przez 100 000-letnie cykle, przeplatane cieplejszymi okresami interglacjalnymi. Potem nastąpiły "epoki lodowcowe" górnego plejstocenu (125-12 Kya), które były też bardzo chłodne okresy zlodowacenia, które były przełamywane przez okresy ciepłe. W chłodniejszych okresach woda byłaby zamknięta i zamarznięta w lodowcach, powodując obniżenie poziomu mórz. Niższe poziomy mórz prawdopodobnie miały ogromny wpływ na mobilność homininów, ponieważ odosobnione wyspy lub odległe kontynenty mogły zostać połączone z lądem za pomocą suchego lądu. Następnie, gdy pokrywy lodowe stopią się w ciepłej fazie interglacjalnej, regiony te mogą ponownie stać się wyspami i izolować żyjące na nich gatunki. Skamieliny zwierząt i roślin w miejscu homininów są przydatne do rekonstrukcji paleośrodowiska w sposób, który różni się od maleńkich otworów w rdzeniach morskich. Na podstawie tego, co wiemy o zróżnicowanej ekologii żyjących gatunków byków (antylop i tym podobnych), typy kopalnych byków na danym stanowisku mogą wskazywać, czy były to otwarte łąki, czy bardziej zamknięte lasy. Kopalne małpy są również wiarygodnymi wskaźnikami środowiska: jeśli mają nadrzewne adaptacje w szkielecie lub zębach, które są związane z jedzeniem liści (jak współczesne colobiny), to prawdopodobnie środowisko było zalesione. Jeśli mają adaptacje ziemskie, takie jak pawiany, miejsce homininów może być znacznie bardziej otwarte, suche i podobne do sawanny. Często zęby krokodyli są porozrzucane wszędzie w miejscach w pobliżu starożytnych wodopojów lub pradawnych koryt rzecznych. Im więcej zwierząt może wykorzystać paleontolog do rekonstrukcji paleośrodowiska, tym silniejsza jest nauka.
Początki Człowieków (23)
GEOLOGIA I METODY DATOWANIA
Przed zorganizowaniem ekspedycji paleontolodzy najpierw patrzą na mapy geologiczne regionu, aby określić, czy rzeczywiście występuje odpowiedni okres czasu. Następnie najlepiej przyjrzeć się zdjęciom lotniczym lub udać się samolotem, samochodem lub pieszo, aby ustalić, czy te skały są odsłonięte i dostępne oraz wolne od gęstej roślinności lub pokrywy wodnej. Często paleontolodzy i geolodzy wspólnie badają interesujące obszary. Jeśli interesują ich skamieniałe homininy, patrzą na skały datowane na miocen, pliocen i plejstocen. Ale jeśli zauważą ogromne kości wynurzające się z kredowych warstw skalnych, zaalarmują swoich kolegów, którzy badają dinozaury. Po odkryciu skamieniałości i artefaktów należy oszacować dla nich dokładniejsze daty, aby umieścić je w kontekście ewolucji człowieka. Albo same skamieliny i artefakty są datowane (datowanie bezpośrednie), albo skały i osady zawierające skamieliny są datowane (datowanie pośrednie). Często najlepiej jest datować skały, a nie skamieniałości lub artefakty, aby uniknąć uszkodzenia okazów. Ze względu na zakresy błędów związane z metodami datowania stosuje się kombinację metod. Łącząc metody względne (przybliżony wiek) z metodami bezwzględnymi (dokładny wiek z zakresami błędu) możliwe jest umieszczenie próbki w bardziej precyzyjnym kontekście. Metody względne mogą jedynie wskazać, czy skamielina jest starsza czy młodsza od czegoś innego. Nie dają one dokładnych dat, ale zasady opierają się na trzech prawach stratygrafii:
1. Prawo superpozycji - depozyty głębiej pod ziemią są starsze niż te, które znajdują się bliżej powierzchni.
2. Prawa Pierwotnej Horyzontalności - warstwy są osadzane (np. przez powietrze, wodę, erupcję wulkanu) poziomo i jeśli nie wydają się poziome, to procesy geologiczne, takie jak uskoki lub kopuły, zmieniły je.
3. Prawo pierwotnej ciągłości ekwiwalentnej osadów, które są oddzielone na przykład kanionem, były wynikiem tego samego zdarzenia depozycji i kiedyś należały do tej samej ciągłej warstwy, zanim erozja je oddzieliła.
Podczas długich okresów historii Ziemi pole magnetyczne było przeciwne do tego, czym jest teraz. Obecny stan nazywa się "normalnym", a przeciwieństwo jest znane jako "odwrócone". Kryształy żelaza w rozgrzanych skałach będą działać jak magnesy i orientują się w kierunku biegunów, gdy skała ostygnie. Tak więc charakter pola magnetycznego Ziemi w czasie, w którym powstała skała, jest zapisywany w jego biegunowości. Skały, które są bezwzględnie datowane i ujawniają swoją polarność, pozwalają geologom na skonstruowanie sekwencji zmian paleomagnetycznych w czasie, które są weryfikowane przez zapis znany z rdzeni głębinowych. Następnie, jeśli skamielina zostanie znaleziona w skale, która nie pozwala na datowanie bezwzględne, geolog może zastosować logikę datowania względnego do odczytu paleomagnetycznego (w oparciu o to, czy skała jest "normalna" czy "odwrócona"), aby umieścić skamieniałość w określonym przedziale czasowym w historii. Paleoantropolodzy znajdują znacznie więcej skamieniałości niehomininów niż homininów, co pozwala im zrekonstruować środowisko, w którym żyli homininy. Skamieniałości niehomininów są również pomocne we datowaniu względnym za pomocą techniki zwanej biostratygrafią. Wiek skamieniałości hominina można określić na podstawie skamieniałości innych dobrze znanych (i dobrze datowanych) zwierząt, które są z nim powiązane. Na przykład, jeśli pewien dobrze znany gatunek skamieniałej świni zostanie znaleziony w tej samej warstwie co skamielina hominina w miejscu A, a ten gatunek świni znajduje się również w miejscu B, datowanym na 6 milionów lat temu, to jest prawdopodobne, że miejsce A ma również 6 milionów lat temu i dlatego skamieniałości homininów również są w tym wieku. Jak w ogóle datowano świnię ze stanowiska B? Datowanie bezwzględne to sposób na oszacowanie dat skamieniałości i artefaktów, zwykle w zakresie lat. Metody datowania radiometrycznego są ważnym elementem podejścia datowania bezwzględnego. Metody datowania radiometrycznego opierają się na znanej stałej szybkości rozpadu izotopów (promieniotwórczych form pierwiastków) poprzez utratę cząstek w jądrze atomów. Datowanie radiowęglowe lub datowanie węglowe jest najbardziej znaną z tych metod. Ponieważ wszystkie żywe organizmy zawierają węgiel, artefakty, takie jak drewno i kości, można bezpośrednio datować tą metodą. Rośliny i zwierzęta pobierają C14, izotop węgla przez całe życie. Kiedy umierają, C14 zaczyna się rozpadać na azot (N14). Śmierć włącza radiometryczny zegar. Rozpad C14 odbywa się w stałym znanym tempie, a kiedy mierzy się C14 obecny w starej kości, można mu określić wiek na podstawie tego, jak bardzo uległ rozkładowi. Czas potrzebny do rozpadu połowy radioaktywnego izotopu w próbce to okres półtrwania. Okresy półtrwania są różne dla każdego izotopu promieniotwórczego. Okres półtrwania dla C14 wynosi 5730 lat, więc w tym czasie połowa próbki C14 uległa rozkładowi. Przy tak krótkim okresie półtrwania próbek powyżej 50 000 lat nie można datować według rozpadu C14, ponieważ w tym czasie cały C14 ulegnie rozkładowi i zniknie z próbki, co uniemożliwi pomiar i datowanie. Za pomocą tej metody datowano skamieniałości homininów z Europy i Australii, ale odpowiedni zakres datowania węglem wyklucza większość zapisów kopalnych dotyczących ewolucji człowieka. Okres półtrwania potasu (K40) wynosi 1,3 miliarda lat. Rozkłada się znacznie wolniej niż C14, więc metody datowania wykorzystujące potas mają zastosowanie do reszty skamieniałości homininów powyżej 100 Kya. Garniss Curtis i Jack Evernden opracowali metodę śledzenia rozpadu K40 na argon (Ar40) w skałach, metodę podobną do datowania węglem, z tą różnicą, że zamiast datować obiekt zainteresowania, datuje się skały wokół obiektu. Podczas ekstremalnego ogrzewania, które występuje podczas erupcji wulkanicznych, argon jest usuwany ze skał wulkanu. Gdy lawa ochładza się, zawiera radioaktywny izotop K40, ale nie zawiera Ar40. Ochłodzenie i zestalenie skały wulkanicznej ustawia zegar na zero i uruchamia go (tak jak wtedy, gdy organizm umiera za datowanie węglem). K40 z czasem rozpada się na stabilny Ar40 w zastygłej skale lawy. Jeśli ta skała zawiera skamieliny hominina lub jeśli znajduje się tuż nad lub tuż pod skamieniałością hominina lub kamiennym narzędziem, próbka zostanie pobrana, a następnie zabrana do laboratorium w celu podgrzania. Naukowcy zmierzą względne ilości K40 i Ar40, aby określić, w jakim stopniu nastąpił rozkład i na tej podstawie mogą oszacować, jak stara jest skała, a zatem jak stara jest powiązana z nią skamielina. Nieco bardziej precyzyjna odmiana metody K/Ar opiera się na tej samej idei i mierzy wielkość rozpadu Ar40 na Ar39, w metodzie zwanej 40Ar/39Ar. Oczywiście randki K/Ar nie działają w miejscach z homininami bez wulkanów w pobliżu, które wypluwają popiół i lawę. Na szczęście wiele skamieniałości homininów w Afryce Wschodniej znajduje się w pobliżu Wielkiej Doliny Ryftowej, która jest wulkanicznie i geologicznie aktywną strefą uskoków, gdzie Afryka dosłownie się rozpada. Wiele wulkanów Riftu było bardzo aktywnych podczas ewolucji homininów, kładąc warstwy popiołu zwane "tufami", które umożliwiają geologom zbudowanie sekwencji datowania. Na przykład skamieniałości zachowane między popiołem A datowanym na 1,6 miliona lat a popiołem B datowanym na 1,5 miliona lat szacuje się na 1,6-1,5 miliona lat temu, czyli średnio na 1,55 miliona lat temu. Tufy są nie tylko wygodne do datowania bezwzględnego, ale ponieważ każdy tuf z erupcji wulkanu ma swój unikalny chemiczny odcisk palca, tufy można śledzić z dużych odległości, aby umożliwić skorelowanie i porównanie miejsc. Miejsca homininów pomiędzy zakresami datowania radiowęglowego i K/Ar są datowane technikami takimi jak elektronowy rezonans spinowy (ESR) i datowanie serią uranu (oparte na rozpadzie izotopu uranu) i działają one najlepiej między 500 a 50 Kya. ESR zależy od mierzalności promieniowania emitowanego podczas rozpadu izotopów. Termoluminescencja (TL) również opiera się na emitowanym promieniowaniu, ale szacuje ilość promieniowania przez podgrzanie skały ("termo") i pomiar ilości emitowanego światła ("luminescencja"). TL jest preferowaną metodą datowania palenisk w bazach mieszkalnych i na kempingach, ponieważ możliwe jest rozszyfrowanie, czy i kiedy skała została podgrzana (to znaczy "podgrzana", ponieważ została po raz pierwszy podgrzana podczas jej formowania). Różne metody datowania są odpowiednie dla różnych materiałów w różnym wieku. W miarę możliwości naukowcy stosują wiele metod. Jednak w przeciwieństwie do datowania radiowęglowego, które czasami pozwala datować artefakt w ciągu 100 lat, metody skamieniałości homininów rzadko zbliżają się do szacowanych zakresów w ciągu kilku tysięcy lat. Wraz z rozwojem technologii rozdzielczość technik datowania staje się coraz bardziej precyzyjna, a zakresy błędów stają się coraz mniejsze. Oczekiwaniem staje się możliwość precyzyjnego określenia miejsca skamieniałości w historii ewolucji homininów (w ciągu kilku tysięcy lat, a nie kilkudziesięciu tysięcy lat).
Początki Człowieków (22)
Prehistoryczne dowody
CO TO JEST Skamieniałość?
Chociaż zwykle trudno je znaleźć, skamieniałości nie są trudne do zobaczenia. To, co jest zaskakujące dla wielu początkujących łowców skamielin, to to, jak wyglądają jak żywe skamieniałe zwierzę lub roślina. Nie ma potrzeby używać wyobraźni, aby dostrzec skamieniałości w ziemi, jakby wyczarowywały na niebie chmury w kształcie zwierząt. Organizmy biologiczne są symetryczne, matematyczne, mają pewne wzory i w większości przypadków ulegają skamieniałości, zachowując znacznie wygląd, jaki miały w życiu, nawet jeśli stają się płaskie i przypominają prehistoryczną śmierć na drodze po milionach lat geologicznego nacisku. W zależności od tego, gdzie się patrzy, skamieniałości są czasami trudniejsze do wyśledzenia niż trofea, ale w kilku wyjątkowych miejscach, takich jak dzikie tereny wokół jeziora Turkana w Kenii, trudno jest przejść bez zmiażdżenia fragmentów skamieniałości pod butami.

Skamieniałości to zmineralizowane szczątki niegdyś żywych organizmów. Mineralizacja materiału organicznego następuje poprzez wymianę wód gruntowych lub poprzez wypłukiwanie minerałów między skamieniałościami a otaczającą ziemią. Chemikalia często zastępują również pierwiastki nieorganiczne w kościach i zębach. Skamieniałości mogą powstawać z takich rzeczy jak kości, zęby, rośliny, drewno, włosy, pióra, odciski stóp (np. dwunożny chodnik w Laetoli w Tanzanii, około 3,6 mln lat temu), ślady, ślady, odciski ciała, kał i wymiociny. Tkanka miękka, podobnie jak skóra, rzadko ulega fosylizacji, ale jest to możliwe i zwykle przybiera formę odcisku lub "skamieniałości śladowej". Kiedy bardzo młode skamieniałości nie zamieniają się całkowicie w skałę, nazywa się je "subfosylnymi". Należą do nich szczątki szkieletowe zwierząt, które niedawno wyginęły, jak wiele gatunków lemurów na Madagaskarze, z których część zaginęła dopiero w ciągu ostatnich 200 lat. Subfosyli nie należy mylić z "żywymi skamieniałościami", czyli organizmami, które niewiele się zmieniły w czasie, jak krokodyle, które wyglądają bardzo podobnie do skamieniałości swoich 80-milionowych przodków. Skamieniałości są często trudne do znalezienia ze względu na niewielką szansę na to, że martwy organizm rzeczywiście zostanie zachowany, a jeszcze mniejsze prawdopodobieństwo, że ktokolwiek znajdzie go miliony lat później
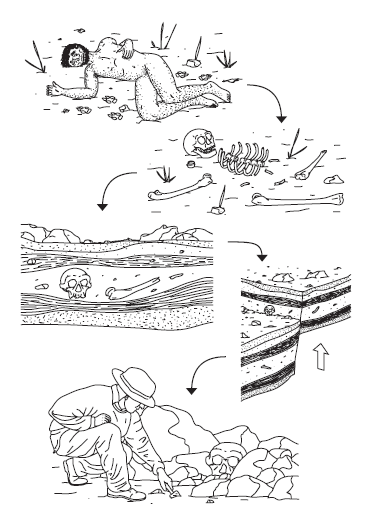
Życie skamieniałości. H. erectus ginie na sawannie i może zostać pożarty lub nie przez padlinożerców, takich jak mięsożercy i ptaki. Następnie, szczególnie jeśli środowisko jest suche, ciało ulegnie rozkładowi, pozostawiając tylko szkielet, a kości mogą zostać rozrzucone przez zwierzęta i żywioły. Z biegiem czasu kości zakopują się naturalnie w osadach niesionych przez wodę lub powietrze. Procesy geochemiczne i ciśnienie pod ziemią zamieniają kość w skałę. Następnie procesy geologiczne, takie jak uskoki (pokazane tutaj) lub prosta erozja, mogą odsłonić skamieliny na powierzchni, aby paleontolog mógł je odkryć. Paleontolog przeprowadziłby wykopy wokół tego miejsca, aby znaleźć jakiekolwiek inne szczątki szkieletu, które, miejmy nadzieję, tam się zachowały.
Jeśli dana osoba chce stać się skamieliną, musi upewnić się, że zostanie szybko pochowana, zanim padlinożercy przeżują i rozrzucą jego kości. Powinni planować umrzeć w pobliżu rzeki, stawu lub jeziora lub wśród wydm, gdzie woda i powietrze stale gromadzą osad. Osad nie może być zbyt kwaśny ani zawierać zbyt wielu mikroorganizmów, ponieważ sprzyjają one degradacji szkieletu. Najlepsze są bardzo suche, suche miejsca, ale pochówek popiołu wulkanicznego jest najlepszym scenariuszem, ponieważ w zależności od geochemii wyrzuconego osadu, osoba może spełnić wymagania, aby stać się skamieniałością, a także mieć możliwość dobrego datowania przez geochronologię. Oczywiście ostatecznym wymogiem stania się skamieliną jest to, aby ktoś faktycznie odkrył, więc ktoś musiałby narysować mapę swojego miejsca pobytu za pomocą symboli, które będą zrozumiałe za milion lat (kiedy współczesne języki rozwiną się nie do poznania), na materiale, który również zostanie zachowany przez eony.
Początki Człowieków (21)
MAŁPY I MAŁPY CZŁEKOKSZTAŁTNE
Małpy dzielą się na grupy geograficzne, Nowy Świat (Ameryki) i Stary Świat (Europa, Azja, Afryka). Ludzie należą do infrarzędu Catarrhini, który obejmuje małpy Starego Świata (Superfamily Cercopithecoidea) i małpy człekokształtne (Superfamily Hominoidea). Catarrhines mają formułę dentystyczną 2:1:2:3, co oznacza, że na ćwiartkę jamy ustnej (są cztery: górna prawa szczęka, górna lewa szczęka, dolna prawa szczęka, dolna lewa szczęka) dorośli mają dwa siekacze, jeden kieł, dwa przedtrzonowce i trzy trzonowce. Pawiany (Papio) i makaki (Macaca) dominują w grupie małp Starego Świata ze swoją różnorodnością i imponującym zasięgiem geograficznym (pawiany rozciągają się od północy do Afryki Południowej, a makaki pokrywają znaczną część południowej, wschodniej i południowo-wschodniej Azji, a nawet docierają do Europy Zachodniej) . Małpy Nowego Świata (infraorder Platyrrhini) mają o jeden przedtrzonowiec więcej na kwadrant niż katar (2:1:3:3), co jest stanem pierwotnym. Mają również chwytne ogony, z których niektóre są pozbawione futerka na spodzie, z odpowiednikiem odcisków palców dla lepszego chwytu. Małpy z Nowego Świata są również bardziej akrobatyczne na drzewach niż małpy ze Starego Świata. Typy małp Nowego Świata obejmują kapucynki, wyje, czepiaki, małpy wiewiórki, marmozety i tamaryny. Żaden żywy naczelny nie je wyłącznie liści, owoców ani owadów, ale to nie zabrania im rozwijania specjalnych adaptacji żywieniowych. Małpy jedzące liście (colobiny) mają długie jelita i wklęsły żołądek do trawienia celulozy, podobnie jak krowy trawią trawę. Ich zęby są również przystosowane do ścinania liści z ostrymi grzebieniami na zębach trzonowych w stanie zwanym bilofodoncją. Colobiny są bardziej siedzące niż inne małpy i mają mniejszy rozmiar mózgu w stosunku do wielkości ciała, czego można się spodziewać, biorąc pod uwagę ich dietę niskiej jakości. Gibbons i siamangi (rodzina Hylobatidae) z Indonezji i Azji Południowej są najbardziej zawieszającymi ze wszystkich naczelnych i są znane jako "mniejsze małpy człekokształtne". Są też najmniej dymorficzni płciowo z człekokształtnych. Poruszają się po drzewach prawie wyłącznie dwurękim ramieniem i czworonożnym wspinaniem się między walkami. Na ziemi chodzą na dwóch nogach, ale jest to rzadkie. Mają bardzo długie ramiona i dłonie oraz bardzo krótkie nogi i stopy (mniejszy ciężar na dole dla lepszego kołysania się przez drzewa). Gibbony i siamangi tworzą monogamiczne pary, w wyniku czego mają niewielki dymorfizm płciowy. Podobnie jak małpy Starego Świata, mają również modzele kulszowe, co jest przystosowaniem do siedzenia, które obejmuje chropowate opuszki na skórze w pobliżu pośladków i pogrubioną kość pod spodem, ale inne małpy nie. Większość diety gibonów to owoce (figi) i mogą one ważyć od 4 do 13 kg w zależności od gatunku, a samce i samice są tej samej wielkości. Dzięki pionierskim pracom terenowym prowadzonym przez prymatologów JaneGoodall (szympansy), Dian Fossey (goryle) i Biruté Galdikas (orangutany), niewiarygodne ludzkie szczegóły dotyczące zachowania wielkich małp i biologii nieustannie zmuszają nas do ponownej oceny tego, co to znaczy być człowiekiem. Te kobiety zanurzyły się w małpim życiu, podchodząc bliżej i osobiście ze stworzeniami i ustanowił precedens dla badaczy i działań ochronnych do naśladowania. "Wielkie małpy człekokształtne" są znacznie większe niż gibony i siamangi i obejmują orangutany (jedyna azjatycka małpa człekokształtna), goryle, szympansy i bonobo. W przeciwieństwie do pomniejszych małp człekokształtnych nie są one monogamiczne, więc ich anatomia odpowiednio się różni. Orangutany (Pongo pygmaeus) są również znane jako małpy człekokształtne i żyją cicho i przeważnie samotnie w zanikających lasach na wyspach Borneo i Sumatra. Ich strategia reprodukcyjna jest znana jako rozproszona poligamia i mają dopasowane ciała dymorficzne płciowo. Samce mogą ważyć do 200 funtów, ale samice ważą mniej niż połowę tego. Samice i ich potomstwo bronią terytoriów przed innymi samicami, podczas gdy samce próbują zapewnić sobie przywileje godowe, kontrolując kilka różnych terytoriów samic. Duże dominujące samce mają szerokie policzki, a także używają worków gardłowych do wydawania głośnych, ryczących pieśni. Orangutany są w większości nadrzewne i jedzą owoce z drzew, ale czasami samce stają się zbyt duże, aby być całkowicie nadrzewnymi i muszą spędzać więcej czasu na ziemi.
Afrykańskie małpy człekokształtne, występujące głównie w regionach zachodniej i środkowej Afryki, są najbardziej lądowymi człekokształtnymi (z wyjątkiem ludzi, którzy są najbardziej ziemskimi człekokształtnymi ze wszystkich). Afrykańskie małpy człekokształtne obejmują Gorilla gorilla (podgatunki obejmują "nizinę zachodnią", "nizinę wschodnią" i bardzo zagrożonego goryla "górskiego") od Kamerunu po góry Wirunga wzdłuż granicy Konga, Rwandy i Ugandy; Pan paniscus (bonobo, dawniej zwany szympansem karłowatym) ze środkowego Konga; oraz Pan troglodytes ("powszechny" szympans) z zachodniej, środkowej i wschodniej Afryki, głównie z Tanzanii, Wybrzeża Kości Słoniowej i Ugandy. Podczas chodzenia na czworonogach wszyscy chodzą na knykciach paliczków dłoni zarówno po ziemi, jak i po poziomych gałęziach drzew. Z największym rozmiarem ciała goryle są najbardziej lądowe. Goryle często żywią się roślinnością na ziemi, w przeciwieństwie do wykorzystywania źródeł pożywienia, takich jak owoce i orzechy na drzewach, takich jak mniejsze szympansy cielesne i bonobo. Jako największe żyjące naczelne samce goryli mogą ważyć do 400 funtów (200 kg), a samice są o połowę mniejsze od samców. Są łagodnymi, towarzyskimi zwierzętami. Jednak szympansy i bonobo są znacznie żywsze. Szympansy są znacznie mniej dymorficzne płciowo z samcami ważącymi około 150 funtów (68 kg), a samice około 120 funtów (56 kg). Bonobo są nazywane szympansami karłowatymi ze względu na ich nieco mniejsze i smukłe ciała w porównaniu do szympansów pospolitych. W dużej mierze ze względu na wielkość ciała goryle gniazdują na ziemi, w przeciwieństwie do szympansów i bonobo, które budują gniazda do spania na drzewach. Poruszając się nadrzewnie, goryle, bonobo i szympansy stosują zarówno czworonożne, jak i zawieszone (wiszące) zachowania lokomotoryczne. Chociaż większość wspinania się goryli charakteryzuje się powolną deliberacją, nawet te największe potrafią wykonywać bardzo szybkie, energiczne wymachy rękami, jak pokazy lub uciekające zachowania szympansów i bonobo. Częstotliwość zachowań nadrzewnych, zwłaszcza u goryli, zależy od wielkości drzew, które je przyjmują. Siedliska goryli górskich i nizinnych są wystarczająco wyraźne, aby uzasadnić różnice anatomiczne między podgatunkami, ponieważ siedliska górskie goryli górskich oferują mniej drzew, po których można się wspinać, w porównaniu z lasami deszczowymi, które zamieszkują goryle nizinne. Strategie rozrodcze goryli i szympansów różnią się. Goryle łączą się w pary w obrębie jednego lub dwóch samców poligamii, w których jeden lub dwa samce mają wyłączny dostęp do grupy samic. Taki system jest korzystny zarówno dla mężczyzn, jak i kobiet, więc termin "harem", który jest piętnem związanym z uciskiem kobiet, nie jest już stosowany do zachowań reprodukcyjnych goryli. Samce czerpią korzyści ze znajomości ojcostwa, a samice korzystają z ochrony samca przed innymi samcami, które mogą zaszkodzić ich potomstwu. Z drugiej strony szympansy mają znacznie bardziej płynną strategię godową. Ich jednostki społeczne nazywane są fuzją rozszczepienia, ponieważ grupy łączą się, a następnie rozpraszają, a następnie regularnie łączą. W takich warunkach kojarzenia mogą mieć miejsce między wieloma różnymi parami męsko-żeńskimi, a hierarchia społeczna w grupach może decydować, kto z kim się kojarzy. Podobnie jak większość naczelnych i wszystkie małpy człekokształtne, potomstwo szympansów pozostaje z matkami, a nie ojcami przez cały okres rozwoju. Wszystkie naczelne wyewoluowały ze wspólnego przodka z rybami i chociaż kuszące jest założenie, że rybacy byli jedynymi ludźmi, wiadomo, że szympansy łowią termity. Mimo to, ludzie mają spory udział w różnicach od swoich najbliższych krewnych szympansów, poza tym, jakie ofiary wybierają do łowienia, a te cechy zostaną omówione w dalszej części.
Początki Człowieków (20)
NACZELNE SSAKI
W przypadku odzieży, języka, samochodów i telefonów komórkowych łatwo zapomnieć, że ludzie to naczelne. Anatomia i genetyka wskazują, że naczelne jako grupa są najściślej spokrewnione ze zwierzętami jeszcze mniej podobnymi do człowieka: colugo (Dermoptera), które są szybującymi ssakami z Azji Południowo-Wschodniej, nazywanymi "latającymi lemurami" i ryjówkami (Scandentia), które są małymi ryjowatopodobnymi zwierzętami, które mieszkają również w Azji Południowo-Wschodniej. Wszystkie naczelne, z kilkoma wyjątkami, dzielą ogólne trendy w zachowaniu, wielkości mózgu, pojedynczym potomstwie (nie w miotach), wydłużonych stadiach wzrostu i rozwoju (tj. wydłużonych "historiach życia"), towarzyskości, anatomii, chwytaniu za ręce i stopy oraz pożytecznym kciukiem, paznokcie zamiast pazurów, oczy skierowane do przodu, widzenie stereoskopowe, uogólniony (tj. wszechstronny) plan ciała, uogólnione zęby, zmienna dieta, kostna obudowa trzech kosteczek usznych ("bula słuchowa") i zamknięte oko gniazdo ("orbita"). Naczelne dzieli się na dwie główne grupy lub podrzędy: Strepsirhines ("mokry nos") obejmuje lemury i lorisy, a Haplorhine ("suchy nos") obejmują wyraki, małpy, małpy człekokształtne i ludzi. Lemury to kotopodobne skaczące (dokładnie "pionowe czepiające się i skaczące"), które żyją tylko na Madagaskarze, dużej wyspie na południowo-wschodnim wybrzeżu Afryki. Lorises to grupa na kontynencie afrykańskim, która zawiera wolno poruszające się potto i skaczące galagos lub "bushbabies". Wyraki są nieco trudne do zdecydowanego zgrupowania z lemurami i lorisami lub z małpami i małpami człekokształtnymi. Mają wspólne cechy pochodne z małpami i małpami człekokształtnymi w wielu różnych kompleksach fizjologicznych i morfologicznych, takich jak oko (tj. światłoczuły dół w siatkówce oka dla ostrego widzenia ("dołek siatkówkowy") i brak odblaskowego grzbietu oka, które poprawia widzenie w nocy, znane również jako "tapetum lucidum", które jest tym samym, co lśniące odbicie psiego oka w fotografii z lampą błyskową). Jednak pomimo swoich cech pochodnych, wyraki pasują do tego samego poziomu przystosowania co lemury i lorysy i czasami grupuje się je w małpiatki - nazwa, która podkreśla prymitywne, nie do końca małpie zachowania lemury, lorysy, i wyraki. Wyraki mają prymitywne cechy, takie jak nie zrośnięta szczęka pośrodku i pazury na niektórych palcach (zamiast paznokci). Ten problem z pewnym pogrupowaniem dwóch głównych podrzędów naczelnych sprowadza się do fundamentalnego problemu z klasyfikacją. Zmuszenie organizmu do dzielenia się tylko na jedną grupę, gdy dzieli cechy z dwiema, zawsze spowoduje problem po prostu ze względu na rozgałęziający się i nakładający się charakter ewolucji. Organizmy, które ewoluują w pobliżu podziału głównych grup, będą przypominać obie grupy. Naczelne są inteligentne i mają duże mózgi w stosunku do rozmiaru ciała. Główne cechy zachowania naczelnych - w tym życie w grupie, granice geograficzne, zasięgi rodzinne, przedłużona więź matka-dziecko, długi okres nauki, przynależności i koalicje (wzmocnione przez uwodzenie) oraz komunikacja (wokalna i niewokalna) - są powiązane z ich inteligencją . Zachowanie i ekologia naczelnych są skorelowane z anatomią. Na przykład dieta naczelnych jest powiązana z kształtem zębów i czaszki, które przetwarzają żywność. Wtedy rozmiar ciała i rozmiar mózgu są ze sobą powiązane i oba zależą od jakości i ilości pożywienia. Adaptacje do różnych typów lokomocji naczelnych są widoczne w klatce piersiowej, kończynach, rękach i stopach oraz ogonie. Naczelne manipulują i oceniają swoje środowisko za pomocą rąk, z widzeniem stereoskopowym i barwnym oraz z ich inteligencją. Wskazówki dotyczące systemów kojarzenia się naczelnych pochodzą z cech dymorficznych płci, takich jak rozmiar kła i ciała. Stopień złożoności społecznej naczelnych może być również skorelowany z rozmiarem mózgu, ponieważ złożone sieci społeczne wymagają złożonych sieci neuronowych, aby je śledzić. Wielkość domowników zależy od wielkości grupy, wielkości poszczególnych zwierząt, dystrybucji pokarmu, wielkości siedliska i konkurencji z innymi grupami (w obrębie tego samego gatunku lub nie). Rodzaje organizacji społecznej mogą przekraczać granice nadrodzin w obrębie naczelnych. Na przykład pottos (rodzaj lorise) i orangutany (wielka małpa człekokształtna) są samotnikami. Wszystkie inne typy organizacji społecznych są grupowe, w których dorośli łączą się w grupy, w tym jeden mężczyzna - grupy wielożeńskie (goryle) i wielomęsko-wielokobiece (pawiany i szympansy).
Początki Człowieków (19)
TAKSONOMIA I KLASYFIKACJA
Kategoryzacja to umiejętność, której używa każdy człowiek, nawet w sprawach wykraczających poza nomenklaturę zoologiczną. Formalny system klasyfikacji organizmów lub taksonomia jest niezbędny, ponieważ zapewnia język, dzięki któremu ludzie mogą współpracować, rozumieć nawzajem swoje wyniki i testować nawzajem swoje hipotezy. Nie wszystkie schematy klasyfikacji zwierząt przekładają się na różne kultury lub wytrzymują próbę czasu. Na przykład klasyfikacja zwierząt w starożytnej chińskiej encyklopedii obejmuje następujące grupy: (a) należące do cesarza, (b) zabalsamowane, (c) tresowane, (d) prosięta ssące, (e) syreny, (f) bajeczne, (g) bezpańskie psy, (h) te, które są objęte tą klasyfikacją, (i) te, które drżą jak szalone, (j) niezliczone, (k) bardzo delikatna szczotka z wielbłądziej sierści, (l) inne, (m) te, które właśnie stłukły wazon z kwiatami, (n) te, które z daleka przypominają muchy (Niebiańskie Emporium Wiedzy Dobroczynnej, do której odwołuje się Borges, 1964). Ze względu na swoje logiczne i naukowe podejście, system klasyfikacji stworzony przez Carolusa Linneusza ma uniwersalne przełożenie. W systemie Linneusza rodzaj i gatunek są zawsze podkreślone lub pisane kursywą, a rodzaj można skrócić tak, że Homo sapiens staje się H. sapiens. W czasie, gdy Linneusz nazwał klasę obejmującą ludzi, publicznie lobbował za korzyściami płynącymi z karmienia piersią, więc wybrał "Mammalia" na cześć gruczołów sutkowych. Jego alternatywną nazwą było równie odpowiednie "Pilosa" dla "włochatych rzeczy". Tak więc Linneusz zdecydował się nazwać włochate zwierzęta w oparciu o cechę, która jest funkcjonalna tylko u połowy gatunków, a u nich funkcjonuje tylko przez ułamek życia. Jednak Linneusz nie wiedział w tamtym czasie, że gruczoły sutkowe wyewoluowały z tych samych typów mieszków włosowych, z których wyewoluowały włosy, więc w wielkim schemacie rzeczy żadna z tych gier nazw nie miała znaczenia. Taksonomia organizmów implikuje relacje, które można wykorzystać do budowania filogenezy lub drzew pokrewieństwa z grupami w zagnieżdżonej hierarchii. Dwa gatunki lub gałęzie filogenezy są ze sobą bliżej spokrewnione niż trzeci, trzy gatunki są ze sobą bliżej spokrewnione niż czwarty i tak dalej. Gatunki są uważane za blisko spokrewnione, jeśli istnieją dowody na ich wspólne pochodzenie w ich fenotypach lub genotypach. Dowody na pokrewieństwo ewolucyjne mają formę wspólnych, pochodnych cech, które ewoluowały od ostatniego wspólnego przodka rozważanego gatunku. Brak ogona jest wspólną, pochodną cechą łączącą ludzi i małpy, z wyłączeniem bardziej spokrewnionych małp. Cechy prymitywne lub przodków, takie jak pięć palców u rąk i nóg, nie są znaczące i nie zawierają żadnych informacji na temat pokrewieństwa ludzi, małp człekokształtnych i małp, ponieważ wszystkie grupy mają pięć palców u rąk i nóg. Zbieżne cechy ewoluują równolegle w różnych rodach od wspólnego przodka, który pierwotnie nie miał tej cechy. Na przykład lot ewoluował zarówno u potomków dinozaurów (ptaki), jak i małych ssaków (nietoperze). Pomimo komplikacji, jakie równoległa ewolucja może wnieść do filogenezy ewolucyjnych, zawsze regułą jest skąpstwo. Zasadę skąpstwa najlepiej podsumowuje zasada "brzytwy Ockhama", która mówi, że najprostsze wyjaśnienie jest zwykle poprawne. W przypadku skąpstwa drzewa filogenetyczne opierają się na najmniejszej liczbie zmian ewolucyjnych i najmniejszej zbieżności. Klasyfikacja jest ostatecznie arbitralna. Głównym problemem związanym z narzuceniem taksonomii na przyrodę jest to, że natura nie jest uporządkowana w zgrabne, małe kategorie. W miarę zbliżania się do korzeni linii, coraz trudniejsze staje się wyraźne umieszczenie gatunków kopalnych na jednej z dwóch lub więcej gałęzi, które rozszczepiły się w czasie ich istnienia. Na przykład im bliżej ostatniego wspólnego przodka ludzi i szympansów (LCA) w zapisie kopalnym, tym trudniej jest z pewnością umieścić skamieliny na szympansach lub ludzkiej gałęzi.
Początki Człowieków (18)
WYJAŚNIENIE EWOLUCJI
Trudno pojąć głęboki czas. Nasze wąskie 100-letnie spojrzenie na wszechświat sprawia, że trudno wyobrazić sobie tysiące, nie mówiąc już o setkach tysięcy, milionach czy miliardach lat. Nasze 100 lat to w najlepszym razie zaledwie 0,00000025% historii Ziemi. Alternatywnie, jeśli historia Ziemi skondensuje się do jednego roku, ludzie przybywają na przyjęcie sylwestrowe z mniej niż 30 minutami wolnego czasu. W tych ograniczeniach trudno jest zaobserwować zmiany ewolucyjne na dużą skalę, takie jak te, które zaszły między rybami a rybakami. Ewolucja może nie ma celu, ale też nie jest przypadkowa. Selekcja działa na dostępnych cechach w obecnych warunkach, nie ma pamięci o przeszłości i nie jest napędzana przez wrodzoną potrzebę doskonalenia gatunku. Mutacja przypadkowa, która wprowadza zmienność genetyczną, może być losowa, ale selekcja nie działa losowo. Selekcja faworyzuje cechy korzystne, a nie przypadkowe. Selekcja może działać tylko z tym, co już istnieje i z tym, co jest rozwojowo wykonalne. Nie ma dalekowzroczności w planowaniu nieuniknionych zmian w środowisku, które zmienią to, co faworyzuje w przyszłych jednostkach w tych nowych warunkach. Dzięki czterem siłom ewolucji (mutacji, dryfowaniu, przepływowi genów i selekcji) dobrze zaprojektowane cechy są wytwarzane przez tysiąclecia prób i błędów. Populacje ewoluują, a nie jednostki. Chociaż termin ewolucja jest często poetycko stosowany do fizycznego lub intelektualnego rozwoju osoby przez całe życie, jednostka nie może ewoluować. Dziecko sześciopalczaste urodzone przez pięciopalczastych rodziców nie jest wynikiem ewolucji, chyba że liczba sześciopalczastych w tej populacji wzrasta w czasie w kolejnych pokoleniach. Scenariusze wspierania wzrostu sześciopalcowości w populacji mogą wynikać z efektu założyciela, z wąskiego gardła populacji lub z selekcji preferującej sześć palców u nóg. Ewolucja nie jest "przetrwaniem najsilniejszych". Herbertowi Spencerowi (1820-1903) przypisuje się połączenie tego hasła z teorią doboru naturalnego Darwina, ale jest ono strasznie mylące. W konsekwencji dobór naturalny jest często błędnie interpretowany jako "tylko silni przetrwają" i, niestety, "silni zabijają słabych". Sprawność fizyczna nie musi mieć nic wspólnego z siłą, a jej osiągnięcie niekoniecznie wymaga przemocy fizycznej. Fitness to po prostu sukces reprodukcyjny. Osobniki z korzystnymi przystosowaniami przetrwają i rozmnażają potomstwo z tą korzystną adaptacją, zwiększając w ten sposób ich przystosowanie. Jeśli musi istnieć hasło na ewolucję, być może "przetrwanie montera" jest bardziej odpowiednie. Sukcesu ewolucyjnego nie mierzy się byciem najlepszym. Wystarczy być nieco bardziej uprzywilejowanym niż inni. Na przykład, w kontinuum osób od niskich do wysokich, osoby z dolnego końca spektrum, niekoniecznie najniższe, są faworyzowane przez selekcję, jeśli ogólny poziom niskiego wzrostu jest korzystny. W następnym pokoleniu geny tych lepiej przystosowanych zwiększają częstotliwość w porównaniu do mniej sprawnych. Niższe osobniki nie eliminują gwałtownie z populacji osobników wysokich. Niskie po prostu przetrwają i prześcigają wysokie. (Można to określić jako "pokonywanie konkurencji", ale to zdanie ma również tendencję do malowania niesprawiedliwego portretu ewolucji "zabij lub daj się zabić".) Jeśli dobór nadal będzie silnie oddziaływał na te niższe, ostatecznie te wyższe będą znikają lub, jeśli są izolowane, a następnie faworyzowane przez selekcję, wysocy będą nadal ewoluować jako oddzielna linia.
Kolokwializmy
Ewolucję często oskarża się, że jest "tylko teorią" z powodu pomieszania zwyczajnych i naukowych zastosowań teorii. Codzienne teorie to przeczucia i często mruczy się je z przymrużeniem oka, by kpić z takich jak Sherlock Holmes. Wręcz przeciwnie, teoria naukowa nie jest zmyślonym schematem. Jest to logicznie spójny model lub ramy opisujące zachowanie określonego zjawiska naturalnego, poparte silnymi dowodami eksperymentalnymi. Jest to systematyczny i sformalizowany wyraz wszystkich wcześniejszych obserwacji. Jest przewidywalny, testowalny i nie został sfałszowany. Chociaż, jak każda idea w nauce, jest podatna na fałszowanie. Teorie naukowe, takie jak ewolucja, poparte wystarczającymi dowodami, są uznawane za fakty. Standardowe wyrażenie, że "wierzy się w ewolucję", jest niefortunną konsekwencją niedociągnięć języka angielskiego. Tworzy antagonistyczną dychotomię między ewolucją a religią, sugerując, że ewolucja jest czymś, w co należy wierzyć, w przeciwieństwie do wiary w bóstwo. Nikt nie mówi, że wierzy w grawitację lub elektryczność, a ewolucję należy odpowiednio traktować.
Początki Człowieków (17)
SPECJACJA
Skumulowana mikroewolucja, czyli zmiany częstości alleli w populacji, prowadzi do makroewolucji, czyli specjacji. W tym sensie zmienność istniejąca w populacji ostatecznie wzrasta, aby stać się zmiennością między populacjami. Ewolucja na poziomie gatunku jest wynikiem kumulacyjnej mikroewolucji. Ryby nie ewoluowały i nie ewoluowały w ludzi, zamiast tego podobny do ryb przodek dał początek wszystkim płazom, gadom, ptakom i ssakom, w tym ludziom. Stopniowe zmiany kumulowały się w czasie w płetwach ryb, aby stać się nośnymi nogami do chodzenia, a reszta to (pre)historia. Specjacja allopatryczna występuje, gdy bariera geograficzna, taka jak rzeka, autostrada lub wyspy oddzielone podnoszącym się poziomem morza, izoluje podzbiór populacji, a siły ewolucji działają niezależnie na dwie populacje. Nagromadzone zmiany prowadzą do niezgodności rozrodczej. Specjacja parapatryczna, błotnisty model specjacji, występuje, gdy podzbiór populacji, który obejmuje duży zasięg geograficzny, w różnych siedliskach, zostaje
odizolowany na podstawie odległości, a selekcja działa w różny sposób na pulę genów w zależności od ich różnych siedlisk. Populacje te mogą być utrzymywane razem jako jeden wysoce zmienny gatunek przez strefy hybrydowe, w których dochodzi do krzyżowania. Pawiany, które są małpami z Afryki Północnej do Południowej, są dobrym przykładem tego zjawiska, ponieważ niektórzy naukowcy grupują je w jeden zróżnicowany gatunek, ale inni postrzegają subpopulacje jako swój własny, indywidualny gatunek. Specjacja sympatryczna jest uważana za rzadki rodzaj specjacji, ponieważ występuje bez granic geograficznych lub fizycznych. Specjacja sympatryczna to formacja dwóch gatunków żyjących w tym samym miejscu. Granice behawioralne, takie jak różnice w wokalizacji lub rytuały zalotów, które zapobiegają krzyżowaniu się w całej puli genów lub przepływowi genów, mogą pozwolić doborowi naturalnemu i doborowi płciowemu działać odmiennie w obrębie populacji i skutkować tym, że subpopulacja faktycznie stanie się odrębnym gatunkiem.
Rodzaje czynników izolujących reprodukcyjnie, które mogą prowadzić do specjacji, obejmują nierozpoznawanie partnerów, różnice behawioralne, preferencje siedliskowe, czas, niezgodność morfologiczną lub mechaniczną podczas kojarzenia, niezapłodnienie jaja i niezdolne do życia potomstwo hybrydowe. Żadna z tych rzeczy nie zapobiega przepływowi genów między populacjami ludzkimi pomimo zróżnicowanej i zróżnicowanej natury gatunku. Gatunki są ostatecznie arbitralnymi kategoriami klasyfikacyjnymi umieszczonymi na ciągłych, nakładających się częściach Drzewa Życia. Istnieje wiele różnych definicji gatunków lub pojęć gatunkowych, które działają dla różnych organizmów w czasie i przestrzeni. Koncepcje morfologiczne i genetyczne gatunków grupują razem organizmy, które posiadają, odpowiednio, podobną anatomię i kody genetyczne. Koncepcja gatunków ekologicznych grupuje te, które mogą mieć tę samą morfologię, ale mają inną niszę. Koncepcja gatunków biologicznych grupuje razem organizmy, które mogą z powodzeniem rozmnażać
żywotne, płodne potomstwo. Trwa debata nad najlepszą koncepcją, którą można zastosować do organizmów kopalnych. Jak na razie nie ma skamieniałości homininów zabitych w akcie krycia. (Chociaż istnieją dinozaury z Mongolii, które skamieniały podczas walki, i istnieje skamieniały ichtiozaur (wymarły gad podobny do delfina), który został pochowany podczas porodu). Jedni mogli się z powodzeniem krzyżować. Dlatego pojęcie gatunku biologicznego, które jest pojęciem najczęściej używanym do grupowania żyjących ssaków, nie ma sensu dla paleontologii homininów. Zamiast tego naukowcy muszą przyjrzeć się zmienności morfologicznej w obrębie żyjących gatunków ludzi, małp i małp człekokształtnych, aby ocenić i przewidzieć wielkość zmienności, jakiej można się spodziewać w obrębie gatunku w zapisie kopalnym homininów.
Inteligentny Projekt (ID)
20 grudnia 2005 r. sędzia John E. Jones orzekł w procesie Kitzmiller kontra Dover, że Dover Board of Education w Pensylwanii nie może nakazać nauczania inteligentnego projektu na lekcjach przedmiotów ścisłych, ponieważ nie jest to nauka. Wyjaśnienia nadprzyrodzone są z definicji poza dziedziną nauki. Jednak zwolennicy inteligentnego projektowania nadal lobbują za włączeniem go do programów nauczania przedmiotów ścisłych w szkołach na całym świecie. Inteligentny Projekt (ID) to idea, która zakłada, że wszechświat jest zbyt złożony, aby mógł ewoluować przez dobór naturalny i musi być zaprojektowany przez inteligentną istotę. ID odwołuje się do ducha amerykańskiej demokracji, fałszywie twierdząc, że w środowisku naukowym toczy się debata na temat ewolucji i że studenci zasługują na wysłuchanie obu stron debaty. Jest to jednak mylące, ponieważ debaty naukowe skupiają się na szczegółach dotyczących natury ewolucji, a nie na tym, czy ona występuje. Poza tym nauka nie jest demokracją. Wszystkim pomysłom nie przypisuje się jednakowej wagi. Utrzymujemy i budujemy na ideach, które zgromadziły dobre dowody naukowe i odrzucamy te, które okazały się nieważne. Nauka i technologia nigdy by się nie rozwinęły, gdyby nie działały w ten sposób. Przykłady nieinteligentnego projektu są liczne, nawet u ludzi, którzy często są uważani za szczytowe osiągnięcie projektu. Ludzkie oko ma martwe punkty i widzi wszystko do góry nogami, więc mózg musi odwrócić wszystko właściwą stroną do góry. Ze względu na anatomiczne zmiany w gardle, które są związane z mową, łatwo jest dostać się do tchawicy ("tchawicy") i zadławić się pokarmem. Połączenie wąskiego kanału rodnego (przystosowanie miednicy do chodzenia w pozycji wyprostowanej) i bardzo dużej głowy rozumnych ludzkich dzieci sprawia, że poród jest trudny i często niebezpieczny. Ze względu na ograniczenia w surowcu, z którym musiał pracować dobór naturalny, niektóre cechy ludzkie wydają się być jerrygowane. Ale pokonujemy je za pomocą innych adaptacji anatomicznych i behawioralnych, a nawet kulturowych innowacji, takich jak aneuver Heimlicha.
Początki Człowieków (16)
WYBÓR SEKSUALNY
Darwin przyznał, że dobór naturalny nie może wyjaśnić wszystkich zmian ewolucyjnych. Nawet jeśli weźmie się pod uwagę współczesne koncepcje mutacji, przepływu genów i dryfu genetycznego, nie są one w stanie łatwo wyjaśnić takich cech, jak nieporęczny ogon pawia, który zmusił Darwina do żartowania, że sam widok tego przyprawia go o mdłości. Aby wyjaśnić zagadkowe rodzaje nadmiernych i potencjalnie upośledzających cech, Darwin wymyślił inny rodzaj doboru, dobór płciowy, w którym samce i/lub samice z populacji wybierają partnerów do godów na podstawie cech, które nie zawsze mają wyraźny związek z dopasowaniem. Dobór płciowy jest kształtowany przez różnicę między mężczyznami i kobietami we wskaźnikach reprodukcji. Samce ssaków mają wyższy wskaźnik niż samice; oznacza to, że mogą wydać więcej potomstwa na jednostkę czasu. Samice muszą czekać na ponowne poczęcie tygodniami lub miesiącami podczas noszenia płodu, ale chwilę po poczęciu samce mogą ponownie zajść w ciążę z inną samicą. Istnieją również różnice między płciami w obrębie gatunku w rodzicielskich strategiach inwestycyjnych. U niektórych gatunków samiec inwestuje niewiele w potomstwo poza początkowym wkładem nasienia, podczas gdy samica jest jedynym opiekunem od poczęcia do dojrzałości potomstwa. Ponieważ samica nosi płód i produkuje mleko, kobieca strategia polega głównie na inwestowaniu w czas, zdrowie, odżywianie i energię. Ponieważ samiec nie nosi płodu ani nie produkuje mleka, strategia samca jest najlepsza, jeśli kojarzy się z wieloma partnerami. Męska strategia jest dalej adaptacyjna, ponieważ męskie ojcostwo jest nieznane bez testów genetycznych. Samce mają alternatywne strategie w gatunkach tworzących monogamiczne wiązania par, jak gibony, gdzie samce w dużej mierze przyczyniają się do potomstwa poprzez ochronę terytorium i pilnowanie partnerów. W systemach z płciami, które mają różne strategie, samice są zazwyczaj płcią wybredną. Dokonują największej inwestycji rodzicielskiej, podejmują największe ryzyko i są źródłem ograniczającym sukces reprodukcyjny samców. Dlatego mężczyźni są płcią konkurencyjną. Konkurują z innymi samcami o dostęp do samic. Z ewolucyjnego punktu widzenia scenariusz ten faworyzuje fenotypy, które dają przewagę płciową samcom, nawet jeśli obniżają ich szanse na przeżycie. Samce odnoszące największe sukcesy to na przykład najbardziej agresywne, najsilniejsze, najpiękniejsze lub mające najdłuższe kły (broń konkurencyjna). Kobiety, które odnoszą największe sukcesy w takim systemie, są najlepszymi wyborcami samców. Część trudności Darwina w pojmowaniu doboru płciowego miała podłoże kulturowe; rozumiał męską konkurencję, ale miał słabsze zrozumienie kobiecego wyboru. Dymorfizm płciowy - różnice w wielkości, kolorze lub anatomii między samcami i samicami gatunku - jest wyolbrzymiony w gatunkach, w których różnice w strategiach godowych są wyolbrzymione, a rywalizacja między partnerami jest wysoka. U małp monogamicznych, takich jak gibon, bardzo trudno jest odróżnić samce i samice bez dokładnego zbadania. Ich rozmiary kłówów i ciała są takie same, a samce mają małe jądra, ponieważ potrzeba wytwarzania obfitych ilości plemników jest zmniejszona w przypadku braku konkurencji z samcami. Szympansy mają jednak znacznie większe jądra, ponieważ konkurują o samice w systemie godowym obejmującym wiele samców i samic. Samce i samice szympansów wykazują różnice w wielkości ciała i wielkości kłów, przy czym samce mają ogólnie większe cechy. W systemie goryli samce i samice wykazują jeszcze większy dymorfizm wielkości ciała niż szympansy. Dominujące samce goryli są znane jako "silverbacks", ponieważ mają srebrne pasma włosów na plecach. Srebrnogrzbiety zwykle mają wyłączny dostęp do grupy samic. Odwrotnie, rozmiar jąder u goryli jest bardzo mały, co może korelować z sukcesem w zapobieganiu kojarzeniu się innych samców z samicami w grupie. Kobiety wybierają partnerów, którzy oferują jej bezpośrednie korzyści, takie jak ochrona, jedzenie i opieka rodzicielska. Mogą również mierzyć wskaźniki sprawności samca, które sygnalizują, czy ma on dobre geny, które zwiększą sprawność jej potomstwa. Wykazano, że takie wskaźniki u ludzi są tak proste, jak jasne oczy, lśniące włosy, świecąca skóra, symetryczna twarz i ciało, umiejętność tańca itd., ale wskaźniki sprawności mogą również wydawać się niepoważne, głupie, a nawet uciążliwe, jak u pawia. ogon. Ewolucję takich cech wyjaśnia zasada upośledzenia Zahaviego, która mówi, że mogą ewoluować cechy, które nie mają żadnej widocznej funkcji poza reklamowaniem sprawności jednostki. Oznacza to, że niektóre cechy po prostu zapewniają środki do przechwalania się, że dana osoba jest tak silna lub tak zdrowa, że może przezwyciężyć upośledzenie. Wskaźniki sprawności, niezależnie od tego, czy są utrudnieniami, czy nie, ewoluują w procesie zwanym niekontrolowaną selekcją, który został opracowany przez R.A. Fisher w 1930 roku. Powiedzmy na przykład, że samica preferuje nos większy niż przeciętny. Będzie kojarzyć się z samcami o dużych nosach i wyda potomstwo o dużych nosach. Jej synowie podniosą średni rozmiar nosa w populacji, a córki będą nosiły jej gen preferowania dużych nosów. Jeśli pozwoli się na kontynuowanie cyklu i jeśli duże nosy są rzeczywiście skorelowane z kondycją, samice będą wybierać coraz większe nosy, zwiększając w ten sposób średni rozmiar nosa w populacji. Dobór niekontrolowany odpowiada za ewolucję cech dymorficznych płci oraz ich preferencje.
Początki Człowieków (15)
SIŁY EWOLUCJI
Dobór naturalny działa na jednostkę, ale ewolucja zachodzi na poziomie populacji. Zmiany w częstości alleli w populacji w czasie określa się jako mikroewolucję, która może zachodzić szybko i jest bezpośrednio obserwowalna. W większej skali makroewolucja jest zmianą długoterminową, charakteryzującą się specjacją, która najczęściej zachodzi w czasie głębokim lub geologicznym. Skumulowane zmiany mikroewolucyjne prowadzą do zmian makroewolucyjnych. Mikroewolucja i makroewolucja leżą w kontinuum zmian od małej do dużej skali, które wynikają z tych samych mechanizmów genetycznych i tych samych czterech sił: mutacji, przepływu genów, dryfu genetycznego i selekcji.
Mutacja
Mutacja losowa dostarcza nowego materiału do selekcji naturalnej. Zmiany w kodzie genetycznym są przekazywane potomstwu, gdy podczas mejozy wystąpią przypadkowe błędy w odczycie i kopiowaniu. Mutacje występują również w wyniku ekspozycji na czynniki zmieniające sekwencję, wywołujące mutacje ("mutageny"), takie jak promieniowanie i chemikalia. Większość mutacji powoduje spontaniczne poronienia, jest szkodliwa lub po prostu neutralna. Korzystne mutacje są rzadkie, ale kiedy pojawiają się w gametach i mogą zostać przekazane potomstwu, mogą mieć duży wpływ na ewolucję. Mutacje punktowe, czyli delecje, substytucje lub addycje nukleotydów, są najczęstszym rodzajem zmiany w kodzie genetycznym. Mutacje mogą również obejmować całe lub częściowe chromosomy. Błędy te występują podczas mejozy, kiedy komórki jajowe lub plemniki otrzymują za dużo lub za mało chromosomów, a następnie ulegają zapłodnieniu. Rezultatem jest często poronienie. Trisomia 21, czyli zespół Downa, jest jednym z niewielu możliwych warunków. Występuje w jednym na tysiąc żywych urodzeń. Osoby mają trzy zamiast dwóch kopii chromosomu 21 i rozwijają unikalny, rozpoznawalny kształt twarzy i głowy, są zwykle mniejsze i mają różne stopnie problemów rozwojowych i zdrowotnych.
Przepływ genów
Allele są wymieniane między populacjami poprzez przepływ genów, który może wystąpić między sąsiednimi populacjami lub gdy populacje są nowe w regionie z powodu rozproszenia lub celowej migracji. Przepływ ludzkich genów jest regulowany przez kulturę, która określa, jak często populacje wchodzą w interakcje i krzyżują się, a także jak poruszają się w przestrzeni i czasie. Przepływ genów miesza allele z różnych puli genów, zapobiegając w ten sposób ich rozdzieleniu na odrębne gatunki.
Dryf genetyczny
Przypadkowa zmiana częstotliwości alleli w czasie to dryf genetyczny, który jest stanem naznaczonym brakiem przepływu genów. Dryf jest spowodowany samym przypadkiem, a wynikiem bardzo silnego dryfu jest albo wygaśnięcie alleli, albo fiksacja. Oznacza to, że allele można całkowicie wyeliminować z puli genów, podczas gdy inne mogą zawładnąć każdym osobnikiem. Dryft jest najczęściej spowodowany przez wąskie gardło populacji lub efekt założycielski i jest najbardziej obserwowalny w małych populacjach. Populacja napotyka wąskie gardło, gdy jej rozmiar zmniejsza się stosunkowo szybko, a następnie ponownie rośnie, na przykład w wyniku głodu, wojny, ludobójstwa lub choroby. Efekt założyciela jest rodzajem wąskiego gardła, które pojawia się, gdy mała podgrupa zostaje odizolowana od populacji i rozpoczyna nową populację poprzez kojarzenie się ze sobą, jak gdyby na przykład mała grupa turystów rozbiła się na niezbadanej bezludnej wyspie. dryfowanie, cechy neutralne, takie jak sześć palców (polidaktylia) lub cechy nieprzystosowawcze, takie jak zespół Ellisa-van Crevelda (zaburzenie, które obejmuje karłowatość kończyn, polidaktylię oraz wady rozwojowe serca i kości i występuje z częstotliwościami wyższymi niż normalne w amiszach Starego Porządku w Pensylwanii) są utrwalane po prostu z powodu przypadku i małej liczebności populacji. Dryf powoduje zmniejszenie zmienności w obrębie populacji lub puli genów.
Wybór
Jako najsilniejsza siła ewolucyjna dobór ma zdolność do zatrzymywania lub eliminowania alleli w populacji. Korzystne rozprzestrzenianie się alleli. Znikają niekorzystne allele. Na przykład nowa mutacja lub neutralna cecha, która już istnieje, staje się korzystna w warunkach środowiskowych i umożliwia jednostkom odnoszenie większych sukcesów niż osobom bez niej. Ludzie są generalistami i mogą rozwijać się w wielu różnych warunkach, ale na wszystkie organizmy wpływają zmieniające się warunki środowiskowe, niektóre są po prostu bardziej wrażliwe niż inni. Selekcja faworyzuje osoby z "najlepszymi" cechami, ale jeśli warunki się zmienią, cechy, które jednego dnia są neutralne lub nawet niekorzystne, mogą być korzystne następnego. Ze względu na zmienność utrzymywaną przez mejozę, recesywne cechy, mutacje i przepływ genów oraz ze względu na przypadkowe zachowanie cech z powodu dryfu genetycznego, neutralne i niepożądane cechy zawsze będą się utrzymywać w populacjach. Jeśli warunki zmienią się radykalnie lub szybko, ta zmienność uratuje gatunek, ponieważ dobór naturalny będzie miał więcej do roboty, zanim ten gatunek wyginie.
Hox geny
Nasze ciała składają się z wzorzystych segmentów kontrolowanych przez odpowiednio wzorowane i segmentowane geny lub geny Hox. Zmiany w liczbie tych powtarzających się części (poprzez mutacje, które powodują duplikację genów, wzmocnienie funkcji genu, usunięcie genu lub utratę funkcji genu) mogą prawdopodobnie wyjaśniać "skoki" ewolucyjne, takie jak przyrost i utrata nóg w różnych liniach (wielorybów). , węże, beznogie jaszczurki) oraz dodanie lub utratę zębów, palców u rąk i nóg. Mutacje wpływające na regulację genów Hox spowodują również innowacje rozwojowe, w tym zmiany w czasie, lokalizacji i ilości ekspresji genów Hox. Ewolucja kręgosłupa ma duże znaczenie dla pochodzenia człowieka. Segmenty kręgosłupa (kręgi) uległy zmianom w trakcie ewolucji naczelnych, z których większość przypisywana jest zmianom w lokomocji. Małpy mają sześć kręgów lędźwiowych (w dolnej części pleców), ale ludzie zwykle mają ich pięć. Ponadto zarówno małpy człekokształtne, jak i ludzie stracili razem ogon (sam koniec kręgosłupa). Prawdopodobnie za te zmiany odpowiedzialne są mutacje w genach Hox.
Początki Człowieków (14)
DNA, CHROMOSOMY, KOMÓRKI I DZIEDZICZENIE
Podstawowy plan komórki zawarty w genomie … działają tak dobrze, że każdy człowiek rozwija się z kilkoma defektami z jednego zapłodnionego jaja w skomplikowany zespół bilionów wyspecjalizowanych komórek, które harmonijnie funkcjonują przez dziesięciolecia w ciągle zmieniającym się środowisku.
-Pollard i Earnshaw, 2002 r.
Osoby lub organizmy są zbudowane z narządów, które są zbudowane z tkanek, które składają się z komórek. Ludzie (i wszystko oprócz bakterii i archeonów) są zbudowani z komórek eukariotycznych, które zawierają jądra (sing. nucleus). Wszystkie typy komórek mieszczą się w dwóch głównych kategoriach: komórki somatyczne i gamety. Komórki dzieli się na te dwie grupy zgodnie z ich różnymi funkcjami, różnymi sposobami replikacji komórek i różnymi ilościami materiału genetycznego w jądrze. Jądro komórki zawiera materiał genetyczny w postaci chromosomów, które są bardzo skondensowanymi, długimi nićmi DNA (kwas dezoksyrybonukleinowy). Każde jądro komórki ludzkiej zawiera 23 pary chromosomów. Istnieją 22 pary chromosomów autosomalnych (44 chromosomy) i jedna para chromosomów płci (2 chromosomy), co daje łącznie 23 pary (46 chromosomów). Kobiety mają dwa chromosomy płci X (XX), a mężczyźni jeden X i jeden Y (XY). Szympansy mają 24 pary chromosomów, a ludzie mają jedną parę mniej, ponieważ dwie z nich połączyły się podczas ewolucji. Gamety zawierają dokładnie połowę dopełnienia chromosomów w komórkach somatycznych. Oznacza to, że zawierają jeden z każdego z 23 chromosomów lub 23 niesparowane chromosomy. Podczas zapłodnienia, kiedy plemnik i komórka jajowa łączą się i tworzą zygotę, każda gameta dostarcza połowę niezbędnego DNA do wytworzenia komórek z pełnym zestawem wszystkich par chromosomów, umożliwiając jej normalny rozwój w zarodek i dalej. Kiedy potomstwo dalej się rozwija, w końcu wytworzy własne gamety, które zawierają połowę jego materiału genetycznego, który zostanie przekazany podczas rozmnażania. Wtedy cykl życia trwa. Komórki somatyczne muszą się dzielić i duplikować, aby rosnąć tkanki podczas rozwoju i utrzymywać tkanki, gdy komórki są stare lub uszkodzone i umierają. Podział komórek somatycznych, czyli mitoza, stale zachodzi w całym ludzkim ciele z szacowaną szybkością 300 milionów duplikacji komórek dziennie, która spowalnia się wraz z wiekiem. W wyniku mitozy powstają dwie kopie komórki macierzystej. Gamety dzielą się, aby umożliwić stworzenie potomstwa. Mejoza, proces podziału gamet, który zachodzi w jajnikach u samic ("oogeneza") i jądrach męskich ("spermatogeneza"), skutkuje powstaniem czterech komórek potomnych. Każda komórka potomna dziedziczy tylko połowę chromosomów komórki rodzicielskiej. U kobiet mejoza występuje u rozwijającego się płodu i do czasu narodzin dziewczynki posiadają wszystkie jaja, które będą miały do końca życia. W przeciwieństwie do mężczyzn, mejoza stale uzupełnia nasienie przez większość życia. Wszystkie komórki jajowe mają chromosom X (ponieważ żeńska komórka rodzicielska ma dwa X), ale plemniki mogą zawierać Y lub X. Dlatego to męski wkład w zygotę determinuje płeć potomstwa, ponieważ jeśli plemnik nosi Chromosom Y zapładnia komórkę jajową, dziecko będzie płci męskiej (XY), a jeśli jest to chromosom X, dziecko będzie płci żeńskiej (XX). Skrzyżowanie chromosomów to zjawisko, które występuje tylko podczas mejozy. DNA rodziców zostaje zamienione, a powstałe chromosomy zawierają mozaikę genów matki i ojca. W konsekwencji chromosom może mieć jeden koniec ojcowski i jeden od strony matki. Potomstwo ma nowe kombinacje lub rekombinację genów zarówno matki, jak i ojca, które tworzą zupełnie nową kombinację, która nie istnieje u żadnego z rodziców. Krzyżowanie i rekombinacja mają kluczowe znaczenie dla zachowania zmienności genetycznej w kolejnych pokoleniach, ponieważ potomstwo nie jest klonem rodziców. DNA jest powszechnie nazywane "kodem genetycznym" lub "sekwencją genetyczną". Cząsteczka DNA składa się z dwóch nici sekwencji DNA skręconych razem jak spiralne schody (zwane "podwójną helisą") ze stopniami wykonanymi z nukleotydów. Dla uproszczenia, nukleotydy te są symbolizowane literami, które "określają" sekwencję DNA: A (adenina), C (cytozyna), T (tymina) i G (guanina). Te nukleotydy lub "pary zasad" łączą się i utrzymują razem dwuniciową cząsteczkę DNA. Adenina wiąże się tylko z tyminą (A-T), a cytozyna wiąże się tylko z guaniną (C-G). Na przykład sekwencja ATAG wiąże się z TATC w drugiej nici. Geny to segmenty DNA lub fragmenty kodu, które są opisane przez te litery. Geny kodują aminokwasy budujące białka. Białka tworzą wszystko w ciele, w tym struktury, płyny, hormony, a nawet enzymy, które katalizują lub umożliwiają reakcje budujące inne białka. Znane są loci chromosomalne (sing. locus) wielu genów i gdy jedna osoba ma inny układ nukleotydów dla tego genu niż inna osoba, mówi się, że ma inne allele lub warianty genu. Na przykład, istnieją dwa allele dla rodzaju woskowiny u ludzi, jeden suchy i jeden mokry, ale jest tylko jeden gen woskowiny (ABCC11). Niektóre geny mają więcej niż dwa allele w populacji, tak jak gen określający grupę krwi, który ma allele A, B i O. Osobnik może nosić tylko dwa allele dla danego genu; ten na chromosomie, który odziedziczyli po matce i ten na chromosomie, który odziedziczyli po ojcu. W wielu przypadkach, ale nie we wszystkich, jeden allel będzie dominujący (A) w stosunku do drugiego, który jest recesywny (a). Dominujący allel (A) będzie wyrażany u osobników homozygotycznych, (AA) lub heterozygotycznych (Aa), gdzie maskuje ekspresję allelu recesywnego. Allel recesywny będzie wyrażany tylko u osobników homozygotycznych dla tego allelu (aa), ponieważ istnieje brak dominującego allelu, który by to zamaskował. W eksperymentach Mendla allel koloru żółtego był dominujący i był wyrażony zarówno u osobników "AA" jak i "Aa", a allel koloru zielonego był recesywny, więc był wyrażany rzadziej, ponieważ mógł występować tylko w roślinach "aa". Poza Mendlem wiemy, że allele mogą być współdominujące (np. cecha sierpowatokrwinkowa), a także mogą różnić się indywidualną ekspresją w zależności od obecności innych powiązanych alleli lub alleli regulujących poziom ich ekspresji. Aby geny były prawidłowo wyrażane, DNA musi zostać prawidłowo odczytane i skopiowane podczas podziału komórki oraz musi zostać prawidłowo odczytane i przetłumaczone podczas budowy białka. Jeśli jakikolwiek nukleotyd zostanie odczytany nieprawidłowo w sekwencji, może zostać wywołany niewłaściwy aminokwas, który może zbudować zupełnie inne białko. Na szczęście w system są wbudowane kontrole i wagi, które zapobiegają katastrofie takich błędów odczytu. Na przykład dla każdego pojedynczego aminokwasu istnieje do czterech różnych (choć bardzo podobnych) układów nukleotydów, które tego wymagają. Ta nadmiarowość w kodzie lub tolerancja błędów to liczba błędów odczytu i tłumaczenia (tj. mutacji), które nie są szkodliwe lub śmiertelne. Nadmiarowość kodu to również sposób, w jaki mutacje w sekwencji mogą pozostać neutralne i nie wyrządzić szkody osobnikowi, ponieważ nowy układ nukleotydów nie zawsze wyraża inne białko.
Początki Człowieków (13)
OD MENDLA DO NOWOCZESNEJ SYNTEZY
Darwin nie był w stanie w pełni zrealizować swojej teorii doboru naturalnego, ponieważ w tym czasie nikt nie rozumiał mechanizmu dziedziczenia, siły stojącej za "pochodzeniem z modyfikacją". Darwin położył podwaliny pod ewolucję, wystawiając hipotezę, że cechy są przekazywane potomstwu, a jeśli cecha jest korzystna, częstość jej występowania w populacji będzie rosła, ponieważ potomstwo z tą cechą przeżyje i będzie się lepiej rozmnażać i przekaże tę cechę swojemu potomstwu i wkrótce. Nagromadzenie wielu takich cech zmieni populację z pierwotnego stanu na inny w kolejnych pokoleniach. Jednak nikt nie potrafił wyjaśnić, w jaki sposób te cechy mogą zostać przeniesione na przyszłe pokolenia. Wchodzi austriacki mnich Gregor Mendel (1822-1884). Wykorzystując rośliny grochu w eksperymentach hodowlanych, opracował brakujący element, nieznany sposób dziedziczenia, który pozwolił teorii ewolucji rozkwitnąć i dorosnąć do obecnego stanu. Mendel pozostawał nieznany biologom ewolucyjnym przez trzydzieści pięć lat po opublikowaniu swojej książki z wynikami. Zanim odkryto geny, koncepcje dziedziczenia obejmowały średniowieczną ideę "preformacji", w której gamety zawierały homunkulusa, czyli maleńkiego, kompletnego człowieka. Następnie koncepcja dziedziczenia zmieszanego, poparta m.in. przez Lamarcka i Darwina, postulowała, aby materiał dziedziczny po rodzicach był zmieszany w potomstwie. Problematycznie ta idea zaprzecza utrzymywaniu zmienności, która wyraźnie występuje w populacjach. Przewiduje, że cechy zostaną zamazane lub rozmyte. Na przykład ciemnoskóra matka i jasnoskóry ojciec zawsze mieszali się, aby mieć dzieci o średniej pigmentacji, a oni z kolei oferowaliby swoim dzieciom tylko średnią pigmentację, co ostatecznie doprowadziło do jednolitego koloru skóry w przyszłych pokoleniach. W rzeczywistości ci rodzice mogą wytworzyć spektrum pigmentacji u swoich dzieci, a oni z kolei mogą mieć dzieci, które wyglądają jak oni sami, jak ich partner, jak ich rodzice lub jak rodzice ich partnera. Te i inne wczesne idee, takie jak pangeneza Darwina, zostały porzucone, gdy eksperymenty Mendla wyszły na światło dzienne. Chociaż to, co odkrył Mendel, jest zbyt uproszczonym poglądem, dziedziczność najłatwiej zrozumieć dzięki jego pracy. Eksperymenty Mendla były naukowe. Śledził liczbę potomstwa i ich fenotypy, które wynikały z różnych hodowanych razem roślin. Wykonywał również dużą liczbę krzyżówek, aby zmniejszyć skutki przypadku, co umożliwiło mu również powtórzenie eksperymentów. Zdecydował się zaobserwować kontrastujące, binarne cechy (np. żółto-zielone i gładkie/pomarszczone), które są łatwe do śledzenia przez pokolenia, ponieważ są to cechy odrębne, w przeciwieństwie do ciągłych, takich jak wzrost lub kolor ludzkiej skóry. Według zasady dobrej nauki jego hipotezy były falsyfikowalne, a następnie przydatne do przewidywania dalszych obserwacji. Na przykład odkrył, że kiedy wyhodował prawdziwe odmiany żółtego i zielonego groszku, stosunek potomstwa prawie zawsze wynosił trzy żółte do jednego zielonego. Tego typu eksperymenty pomogły mu w zrozumieniu kilku prostych zasad prostego dziedziczenia, ujętych tutaj w nowoczesne terminy oparte na obecnym rozumieniu (np. termin "gen" został wprowadzony dopiero w 1909 r.).
1. Dwa warianty genów lub allele, po jednym od każdego rodzica, determinują fenotyp. Fenotyp będzie wytwarzany przez dominujący allel, a nie recesywny. Allele nie są mieszane. Są dziedziczone i wyrażane osobno.
2. Każdy rodzic ma dwa allele dla każdej cechy, a szanse są równie prawdopodobne, że potomstwo otrzyma jeden allel od rodzica. Pojęcie to jest znane jako "prawo segregacji" Mendla. W momencie poczęcia, kiedy zygota powstaje z połączenia plemnika i komórki jajowej, allele matki i ojca rozdzielają się losowo na komórki potomne, więc istnieje przewidywalna 50% szansa na otrzymanie jednego allelu lub drugiego. Proces ten przyczynia się do utrzymania zmienności w populacji, na którą oddziałuje dobór naturalny, ponieważ żadne dwa potomstwo nie będzie miało tej samej kombinacji alleli.
3. Cechy są dziedziczone niezależnie. Oznacza to, że na kolor grochu nie miało wpływu to, czy groch był gładki czy pomarszczony. Jest to znane jako "Prawo Niezależnego Asortymentu" Mendla i jest kolejnym sposobem utrwalania zmienności populacji.
Chociaż wyjaśnienia Mendla dotyczące dziedziczenia (oparte na starannie dobranych cechach) okazały się zbyt uproszczone, Mendel był pionierem, który położył podwaliny pod całą współczesną genetykę. Jest powszechnie i szalenie oczywiste, że dziedziczenie w stylu mendlowskim jest raczej wyjątkiem niż regułą. Dziś powszechnie wiadomo, że większość cech jest złożona i wyrażana przez wiele genów, które również są często powiązane w dziedziczeniu. Aby zrozumieć złożone sposoby dziedziczenia większości cech, konieczne jest głębokie zrozumienie współczesnej biologii rozwoju, komórki i biologii molekularnej. Mendel opublikował swoje odkrycia w 1866 roku, zaledwie sześć lat po traktacie Darwina. Został on niezauważony przez społeczność naukową aż do 1900 roku, kiedy wreszcie mechanizm dziedziczenia mógł zostać powiązany z doborem naturalnym, a wkrótce potem narodziła się teoretyczna rewolucja znana jako "Nowoczesna Synteza". Następnie, gdy James Watson, Francis Crick i Rosalind Franklin w 1953 r. stworzyli pierwszy dokładny model cząsteczki DNA, ujawniono fundamentalny składnik genetyczny ewolucji.
Początki Człowieków (12)
NATURALNA SELEKCJA
Chociaż kojarzymy ewolucję z Karolem Darwinem, nie był on pierwszym, który zdał sobie sprawę, że ewolucja zachodzi, ani nie był pierwszym, który szukał mechanizmu. Jean Baptiste Lamarck (1744-1829) dokonał pierwszych solidnych, naukowych prób wyjaśnienia Drzewa Życia. Znaczna część myślenia Darwina była pod wpływem pism Lamarcka i jego poglądów na dziedziczenie "nabytych cech". Klasycznym przykładem ewolucji Lamarcka jest wyjaśnienie ewolucji długiej szyi żyrafy. Zgodnie z tym poglądem żyrafy miały pierwotnie krótkie szyje i za życia wyciągały się, by dosięgnąć wiszących wysoko na drzewach liści. Ich szyje stały się nieco dłuższe, niż gdyby nie zjadały tych niedostępnych liści. Potomstwo proto-żyraf rozciągających szyję odziedziczyło te nieco dłuższe szyje i tak dalej, aż po wielu pokoleniach powstała współczesna żyrafa z długą szyją. Wyobraź sobie w kategoriach ludzkich, łatwiej jest dostrzec błąd w ewolucji Lamarcka. Kobieta, która podnosi ciężary przez całe życie i buduje mięśnie, nie urodzi dzieci z mięśniami, tak jak żyrafy, które wyciągnęły szyję, nie będą miały potomstwa o dłuższej szyi. Wpływy środowiskowe nie są przekazywane przyszłym pokoleniom, więc niestety dla Lamarcka, bez znajomości genetyki i dziedziczenia w tamtych czasach, jego hipoteza nie sprawdziła się. Chociaż nie pierwszy, który rozpoznał ewolucję, Karol Darwin był pierwszym, który wymyślił możliwy do tego mechanizm. Ściśle mówiąc, Charles Darwin i Alfred Russel Wallace (1823-1913) niezależnie opracowali teorię doboru naturalnego, ale Darwinowi przypisuje się większość zasług, ponieważ opublikował poczytną książkę. W O powstawaniu gatunków (1859) Darwin zaproponował mechanizm ewolucji, który - chociaż jest podstawą współczesnej nauki o ewolucji - po prostu otworzył powierzchnię dla odkryć, które miałyby poprzeć ewolucję. Oprócz dowodów, które zebrał podczas podróży H.M.S. Beagle, idee Darwina były również pod wpływem prac ekonomisty Thomasa Malthusa (1766-1834). W swoim Eseju o zasadzie populacji Malthus opisał, jak wzrost populacji ludzkiej jest wykładniczy lub geometryczny, podczas gdy zapasy żywności mogą rosnąć tylko w stosunkowo stabilnym, liniowym lub arytmetycznym tempie. Populacje stale rosłyby przez wieczność, gdyby ograniczanie zasobów, takich jak żywność, nie trzymało ich w ryzach. Szybko rosnąca populacja, która rozmnaża się zbyt szybko ze względu na jej zasoby, może prześcignąć te zasoby, a następnie doświadczyć krachu, dopóki wielkość populacji nie osiągnie poziomu, który zasoby są w stanie obsłużyć. Na wstępie swojej teorii doboru naturalnego Darwin postulował, co następuje: (1) Zdolność populacji do ekspansji jest nieskończona. Ale zdolność każdego środowiska do utrzymania populacji jest ograniczona. Nazwał to "walką o byt" i "gospodarką natury"; (2) Organizmy w populacjach różnią się i ta zmienność wpływa na zdolność jednostki do przetrwania i rozmnażania; (3) Odmiana jest przekazywana z rodzica na potomstwo. Darwin nazwał to "zejściem z modyfikacją", ale w tamtym czasie nie był jeszcze znany mechanizm dziedziczenia. Ujmując to nowocześnie, teorię doboru naturalnego Darwina można podzielić na cztery części.
Zmiana
Organizmy są różne. Każda osoba ma swój własny fenotyp lub zestaw cech fizycznych, takich jak kolor oczu, długość nóg i ton głosu. Cechy fenotypowe obejmują morfologię (takie jak anatomia, rozmiar, kształt, kolor i tekstura), fizjologię i zachowanie. Fenotypy są tworzone zarówno przez naturę (genetyka), jak i wychowanie (wpływy środowiska, takie jak odżywianie podczas rozwoju i czynności podczas życia).
Dziedziczność
Pewna odmiana jest dziedziczona po rodzicach. Potomstwo przypomina rodziców bardziej niż innych, zarówno pod względem wyglądu, jak i zachowania. Genotyp lub skład genetyczny osobnika jest powiązany z fenotypem, a zatem wpływa na niego. Mutacja lub zmiana w DNA osobnika może być korzystna lub szkodliwa, a jeśli wystąpi w gamecie (jajo lub plemniku), może zostać przekazana potomstwu.
Sprawność różnicowa
Sprawność w kategoriach ewolucyjnych nie jest sprawnością fizyczną, taką jak napięcie mięśniowe i zdrowie. Sprawność to liczba potomstwa, które dana osoba produkuje, która jest odpowiednikiem jej sukcesu reprodukcyjnego. Niektóre odmiany są korzystniejsze od innych. Organizmy o korzystnych cechach przetrwają i będą miały więcej potomstwa, które będzie nosiło te korzystne cechy i przekazywało je swojemu potomstwu. Rozmnażanie różnicowe występuje, gdy osobniki o korzystnych cechach rozmnażają się więcej niż osoby bez tych cech. Zwiększone wskaźniki przeżycia i reprodukcji potomstwa danej osoby przyczyniają się do tego, że sprawność tej osoby jest większa niż u innej. Zjawisko to występuje, ponieważ cecha jest "wybierana do" lub "faworyzowana" przez dobór naturalny.
Dostosowanie
Presja środowiskowa, taka jak to opisał Malthus, określi które cechy są bardziej korzystne niż inne. Cechy związane z lepszą reakcją na presję są adaptacyjne i stają się uzasadnionymi adaptacjami, gdy ich częstotliwość wzrasta tak bardzo, że cechę tę posiada większość populacji. Adaptacje mają miejsce, gdy kolejne pokolenia osobników z cechą adaptacyjną osiągają wzrost sukcesu lub sprawności reprodukcyjnej. Na przykład żyrafy, które mają długie szyje, mogą być jedynymi, które mogą sięgać i zjadać liście na najwyższych gałęziach drzew w czasie suszy. Te osobniki o długich szyjach będą miały potomstwo, które ma długą szyję, które jest skuteczne również w czasie suszy i tak dalej. Adaptacje mogą powstać z nieoczekiwanych miejsc. Na przykład posiadanie długiej szyi do jedzenia liści poza zasięgiem może w nowej sytuacji przystosować się do czegoś zupełnie innego, jak trzymanie głowy nad wodą podczas powodzi. W tym hipotetycznym przypadku posiadanie długiej szyi do karmienia jest eksaptycją za pływanie. Postawiono hipotezę, że ludzka krtań jest eksaptacją, ponieważ mogła zostać dokooptowana do wydawania dźwięków mowy po tym, jak już opadła do gardła z innych powodów. Z pewnością stan braku cechy również może być adaptacją. Utrata tylnych nóg u wielorybów doprowadziła do ich opływowych ciał do pływania. Nie wiadomo jeszcze, czy utrata małpy i ludzkiego ogona była zaletą, czy była po prostu wynikiem przypadku. W końcu posiadanie ogona jest korzystne dla balansowania na gałęziach drzew, a jako konsekwencja bycia bezogonowymi małpy człekokształtne nie poruszają się w taki sam sposób jak małpy z ogonami.
Dodatek ludzki
U ludzi niewielki wyrostek przypuszczalnie nie jest funkcjonalny ani niezbędny do przepuszczania pokarmu przez układ pokarmowy. Jest mniejszy niż u innych ssaków, ale wyrostek robaczkowy człowieka ma tendencję do pękania i jeśli nie zostanie szybko usunięty, człowiek może umrzeć. Można się więc zastanawiać, dlaczego wyrostek nie zniknął całkowicie. Może istnieć funkcja, która jest jeszcze nieznana, na przykład podczas rozwoju w macicy. Być może jednak dodatek nie jest na tyle niekorzystny, aby dobór działał przeciwko niemu i usuwał go z fenotypu człowieka. Niewiele osób umiera z powodu pękniętego wyrostka robaczkowego. I wielu z tych, którzy już rozmnażali się i przekazali swoje geny, które zawierają plany budowy wyrostka robaczkowego u ich potomstwa.
Początki Człowieków (11)
Homologia i analogia
Związek między formą a funkcją u zwierząt ujawnia, w jaki sposób pochodzenie lub wspólne pochodzenie wpływa na formy zwierząt. Na przykład kończyny przednie człowieka, wieloryba, psa, konia i nietoperza są podobne pomimo ich różnych funkcji w zakresie manipulacji, pływania, biegania i latania. Te strukturalne podobieństwa lub homologie wynikają z ich wspólnego, choć odległego pochodzenia ssaków i utrzymują się pomimo ewolucji różnych funkcji, które zmieniają je w czasie. Odwrotnie, skrzydła nietoperzy i ptaków reprezentują analogie. Mimo podobnej funkcji struktury nie są równoważne. Skrzydła nietoperza wyewoluowały ze zwykłych czworonożnych przednich kończyn ssaków, a upierzone skrzydła ptaków wyewoluowały z przednich kończyn dwunożnych dinozaurów. Podobne funkcje wyewoluowały niezależnie w tak zwanej ewolucji konwergentnej lub konwergencji adaptacyjnej i nie wskazują na niedawne pokrewieństwo lub dziedziczenie po wspólnym przodku. Innym przykładem są nogi ptaka i nogi człowieka: obie pozwalają zwierzęciu chodzić na dwóch nogach, ale anatomia ich łap jest ułożona zupełnie inaczej. U wielu ptaków pierwszy palec u nogi (a czasami także drugi) jest skierowany do tyłu, aby zachować równowagę i zdolność chwytania, ale duży palec u człowieka wystaje przed innymi. Zwierzęta o podobnej budowie są bliżej spokrewnione pomimo pozornie ogromnych różnic w modyfikacji i wykorzystaniu tych struktur (np. nietoperze i ludzie) niż wiele zwierząt o podobnych funkcjach (np. nietoperze i ptaki).
Cechy szczątkowe
Małe, niefunkcjonalne trzecie sutki nie są niczym niezwykłym u ludzi. Tego rodzaju ewolucyjne resztki, takie jak rzadkie występowanie ludzkiego ogona, są znane jako cechy szczątkowe lub atawizmy. Jako przejaw ewolucyjnej historii organizmu, szczątkowe cechy są pieczęciami w paszporcie ewolucji. Niektóre ssaki, takie jak myszy i psy, mają wiele gruczołów sutkowych do karmienia młodych miotów. Ludzie, podobnie jak większość naczelnych (ale istnieją wyjątkowe strepsiryny z wieloma sutkami), zwykle rodzą jedno dziecko na raz, które wymaga tylko pary gruczołów sutkowych do karmienia. Dodatkowe sutki u ludzi symbolizują wspólne pochodzenie z innymi ssakami, które mają więcej niż dwa sutki. Sutki u samców ssaków nie są cechami szczątkowymi, ponieważ nie sygnalizują odwiecznego stanu produkcji mleka u samców. Męskie sutki są po prostu wynikiem ciekawego procesu rozwojowego. Wszyscy ludzie zaczynają rozwijać się jako samice i dopiero około siódmego tygodnia embrion z chromosomem Y zaczyna rozwijać się jako samiec. To przejście następuje po rozwoju skóry na klatce piersiowej, ale przed rozwojem gruczołów sutkowych pod nią. Męskie sutki są po prostu stemplami w paszporcie rozwoju ssaków
Embriologia
Zrozumienie procesów rozwojowych zarodków ma kluczowe znaczenie dla zrozumienia ewolucji i relacji między organizmami. Bliżej spokrewnione gatunki mają bardziej podobne stadia rozwojowe i procesy rozwojowe. Młode ludzkie embriony przypominają ryby i faktycznie mają struktury przypominające skrzela. U ryb struktury te rozwijają się w skrzela, ale u ludzi rozwijają się one w ucho, szczękę i szyję. Ernst Haeckel opracował hipotezę dotyczącą rozwoju zarodków kręgowców, która została powszechnie zaakceptowana przez wybitnych naukowców początku XX wieku. Nazywano to "ontogeneza rekapituluje filogenezę", co oznacza, że wzrost i rozwój powtarza historię ewolucyjną. Jego hipoteza sugerowała, że podczas rozwoju organizm doświadcza wszystkich etapów swojej historii ewolucyjnej, aż do osiągnięcia pełnego rozwoju. W tym scenariuszu ludzie najpierw rozwijają się w ryby, potem w płazy, potem w gady, potem w ssaki przypominające gryzonie, a w końcu w niemowlęta. Hipoteza Haeckela nie jest już akceptowana, ale jego ogólne obserwacje wciąż brzmią prawdziwie: najwcześniejsze etapy rozwoju w macicy (tj. w łonie matki) przypominają prymitywne pochodzenie, a późniejsze etapy przypominają nowsze wspólne pochodzenie.
Początki Człowieków (10)
Biogeografia
Żeglując po Ameryce Południowej, a ostatecznie po całym świecie, Darwin był świadkiem gatunków zwierząt i roślin, które widziało niewielu Europejczyków, a wiele z nich było nieznanych europejskim naukowcom. Zauważył, że nie tylko wydają się istnieć regionalne podobieństwa między organizmami, ale że organizmy w sąsiednich środowiskach raczej przypominają się nawzajem niż organizmy w podobnych środowiskach w innych częściach świata. Innymi słowy, organizmy ewoluują lokalnie. Słynnym przykładem tego biogeograficznego fenomenu, odnotowanego niegdyś przez Darwina, jest różnorodność zięb na Wyspach Galapagos, odizolowanym archipelagu utworzonym przez wulkanizm i charakteryzującym się suchym, skalistym terenem. Łuszczaki na Galapagos bardziej przypominają zięby na bujnym zachodnim wybrzeżu Ameryki Południowej niż ptaki żyjące w innych surowych, suchych środowiskach. Ponieważ na wyspach brakuje całych grup zwierząt, przybysze mają możliwość wypełnienia otwartych nisz, które często są już wypełnione w ich pierwotnym środowisku. Pierwotne zięby z Galapagos rywalizowały z niewielką liczbą zwierząt na tych stosunkowo odizolowanych wyspach i były w stanie promieniować na wiele gatunków. Wyewoluowały różne rozmiary i kształty dziobów, rozmiary i kształty ciała oraz zachowania żywieniowe. Istnieją zięby, które zachowują się i wyglądają jak dzięcioły, ale ich geny łączą je z innymi ziębami z Galapagos, a nie z prawdziwymi dzięciołami gdzie indziej. Podobnie jak u ludzi, z analiz genetycznych wynika, że Melanezyjczycy o ciemno ubarwionej skórze są bardziej spokrewnieni z Azjatami w tym regionie świata niż z ciemno ubarwionymi Afrykanami.
Skamieniałości i geologia
Od czasów Lyella powszechnie wiadomo, że organizmy wymierają i wymarły w przeszłości, a czasami pozostawiają ślady swojego wcześniejszego istnienia w postaci skamieniałości. Najgłębsze skamieniałości są najstarsze i zawierają cechy łączące ich jako przodków z późniejszymi gatunkami. Zapis kopalny naczelnych to tylko jeden wspaniały przykład. Najwcześniejsze naczelne znajdują się w starszych skałach (65 milionów lat temu) niż pierwsze skamieniałe małpy, które znajdują się w starszych skałach (35 milionów lat temu) niż skamieniałe małpy (20 milionów lat temu). Tylko w skałach, które powstały w ciągu ostatnich 6 milionów lat, paleontolodzy znajdują wyprostowanych przodków współczesnego człowieka. Z biegiem czasu, wraz ze wzrostem cech dwunożności i innych cech ludzkich, takich jak duże mózgi, te małpopodobne maleją.
Sztuczna selekcja
Jako miłośnik hodowli gołębi, Darwin wykorzystał dowody z ludzkiego udomowienia zwierząt, aby poprzeć swoją teorię doboru naturalnego. Od tysięcy lat ludzie hodują selektywnie zwierzęta i rośliny, faworyzując najsmaczniejsze, najbardziej kolorowe, najbardziej wełniane, najmądrzejsze, najserdeczniejsze lub najszybciej biegające. Darwin nazwał hodowlę zwierząt i rolnictwo "sztuczną selekcją" ze względu na to, jak dobrze ludzie potrafią naśladować dobór naturalny. W wyniku sztucznej selekcji powstają nowe odmiany i nowe gatunki. Psy w całej swojej wspaniałej różnorodności są produktem sztucznej selekcji jednego z przodków gatunku wilka. Ludzie niekoniecznie wybierają te same cechy, które sprzyjałby doborowi naturalnemu. Przeżycie można zwiększyć przez interwencję człowieka, dzięki czemu cechy, które potencjalnie mogłyby zaszkodzić powodzeniu organizmu na wolności, mogą się rozwijać. Na przykład arbuzy bez pestek nie są w stanie się rozmnażać, a niektóre wymyślne rasy psów wymagają częstej opieki medycznej ze względu na ich urocze, ale zagrażające zdrowiu cechy, takie jak zbyt krótkie nogi i zbyt krótkie pyski.
Początki Człowieków (9)
DOWODY NA EWOLUCJĘ
Każdy na Ziemi jest związany ze wszystkim, co żyje teraz i co kiedykolwiek żyło w przeszłości. Czternaście miliardów lat temu Wielki Wybuch uformował nasz wszechświat, a dziesięć miliardów lat później materia połączyła się, tworząc planetę Ziemię. Wkrótce potem powstały pierwsze białka, prawdopodobnie w kominach termicznych głęboko w oceanie. Potem rozkwitły prokarionty (organizmy jednokomórkowe), a następnie eukariota (organizmy wielokomórkowe). Zanim byliśmy świadkami skamieniałości "wybuchu kambryjskiego", w szczególności z łupków Burgess w Kolumbii Brytyjskiej, pojawiło się promieniowanie zwierząt morskich, z których wiele przypominało współczesne skorupiaki i owady. Z tych zwierząt wyewoluowały pierwsze zwierzęta z kręgosłupami, strunowce i kręgowce. Ryby płetwiaste dały początek pierwszym czworonogom, którym udało się wyżyć na lądzie do 385 milionów lat temu. Następnie starożytne gady dały początek dinozaurom, ptakom i ssakom. Od jednych z najwcześniejszych ssaków powstała grupa, do której należymy, naczelne. Jak pokazuje Tabela, dobór naturalny powoli i nieświadomie gromadził ludzi, akumulując cechy przez miliony lat ewolucji. Karol Darwin pojął to pojęcie na długo przed pojawieniem się wielu skamielin, które by je potwierdzały, i na długo przed zrozumieniem jakiegokolwiek prawdopodobnego sposobu dziedziczenia tych cech. Podczas swojej pięcioletniej podróży na H.M.S. Beagle (1831-1836), Charles Darwin (1809-1882) zebrał dowody, które wykorzystał do skonstruowania swojej teorii doboru naturalnego. Zbierał zwierzęta, rośliny i skamieniałości oraz kultywował idee, których użył do sformułowania mechanizmu ewolucji i doboru naturalnego. Patrząc wstecz przez współczesną soczewkę, dowody, które obserwował Darwin, są dzisiaj przytłaczająco mocne.
Cecha: Miliony lat temu
Pisanie: 0,006
Rolnictwo: 0,01
Język: 0.1
Symbole: 0,1
Nowoczesny rozmiar mózgu: 0,5
Jedzenie mięsa: 2,5
Narzędzia kamienne: 2,6
Małe kły: 3,2
Dwunożność: 5
Wizja trójkolorowa: 23
Wzór dentystyczny: 35
3 kosteczki ucha: 130
Włosy: 150
5 palców: 340
4 kończyny: 385
Szczęki: 460
3 kanały półokrągłe: 470
Kręgosłup : 520
2 oczy: 550
Początki Człowieków (8)
BIEŻĄCE PROBLEMY
Paleoantropolodzy przeszłości i dnia dzisiejszego odkryli i nadal odkrywaj miejsca w całym Starym Świecie od Syberii po Australię, od Anglii po RPA i od Chin po Portugalię. Dopiero stosunkowo niedawno uczeni wywodzący się z Afryki, Azji, Eurazji, Indonezji i Australii dołączyli do czołówki nauki o pochodzeniu człowieka i jest pewne, że krótka historia filozofii początków człowieka z ich punktu widzenia poszłaby innymi drogami niż ta zakorzeniona w Zachodnia Europa. Wielu naukowców zajmujących się obecnie pochodzeniem i ewolucją człowieka to tylko pierwsze i drugie pokolenie intelektualnego potomka pionierów w tej dziedzinie. Nowe znaleziska skamieniałości, nowe i ulepszone metody datowania oraz nowe analizy genetyczne nieustannie kształtują i często obalają stare idee
1856 Pierwsza małpa z gatunku Dryopithecus-Fossil z Europy
1856 Pierwszy neandertalczyk - Dolina Neander, Niemcy
1868 Pierwsza anatomicznie współczesna skamielina człowieka - Cro-Magnon, Francja
1891 Pierwszy Homo erectus - Jawa, Indonezja
1924 Pierwszy australopitek, "Dziecko z Taung" - Sterkfontein, Republika Południowej Afryki
1959 "Zinj" - czaszka Paranthropus boisei, wąwóz Olduvai, Tanzania
1960 Jane Goodall obserwuje, jak szympansy wytwarzają i używają narzędzi na wolności
1961 Pierwsze zastosowanie datowania potasowo-argonowego w miejscu hominina - Wąwóz Olduvai
1967 Teoria zegara molekularnego zostaje zastosowana w badaniach ewolucji człowieka
1974 "Lucy" - szkielet Australopithecus afarensis
1985 "Chłopiec z Nariokotome" - szkielet Homo erectus
1995 Pierwszy Ardipithecus - najstarszy potwierdzony dwunożny, Aramis, Etiopia
1997 Analiza pierwszego starożytnego DNA skamieniałego hominina (neandertalczyka)
1999 W Bouri w Etiopii odkryto najstarsze kości zmasakrowane narzędziami kamiennymi
2000 Orrorin- Najstarszy rzekomo dwunożny, Tugen Hills, Kenia
2002 Sahelanthropus - Najstarszy hominin, Toros-Menalla, Czad
2000 Zsekwencjonowano ludzki genom
2004 Homo floresiensis - Tak zwani "hobbici" Indonezji
2005 Sekwencjonowanie genomu szympansa
2005 Tylko znany kopalny szympans, Kenia
Teraz, kiedy paleontolodzy dysponują sporą skamieniałością homininów, lepiej radzą sobie z dostrzeganiem najbardziej fragmentarycznych w błocie lub tych niewłaściwie oznaczonych w muzealnych szufladach. A teraz zamiast zaledwie garstki skamieniałych przodków człowieka istnieją dosłownie tysiące okazów co najmniej osiemnastu różnych gatunków.
Dzisiejsi paleoantropolodzy nie są tak skoncentrowani na przejściowych ludziach-małpach, jak w przeszłości. Ponieważ większość drzewa genealogicznego homininów została wypełniona, a zegary molekularne (genetyczne) przewidziały wiek podziału linii ludzkich i szympansów, priorytetem jest znalezienie najwcześniejszych homininów i najwcześniejszych szympansów między 8 a 5 milionów lat temu. Dzięki powiększającemu się zapisowi kopalnemu badanie początków i ewolucji człowieka może odbywać się na wielu etapach prehistorii, a pole to rozszerzyło się znacznie dalej niż to, o czym rozważali Darwin, Huxley i ich kohorta 150 lat temu. Pytania w tabeli 2 podkreślają tylko kilka popularnych tematów badawczych w tej dziedzinie.
• Czy Homo floresiensis jest prawidłowym gatunkiem hominina karłowatego, czy też jest chory?
• Czy współcześni ludzie w Starym Świecie ewoluowali z wcześniejszych form homininów, czy też ewoluowali w Afryce i rozprzestrzenili się na inne regiony, zastępując te formy?
• Czy neandertalczycy byli bezpośrednio zaangażowani w naszą własną ewolucję? Czy współcześni ludzie i neandertalczycy wchodzili w interakcje?
• Dlaczego wyginęli neandertalczycy?
• Dlaczego silne australopity - rodzaj Paranthropus - wyginęły?
• Czy bylibyśmy w stanie rozpoznać najwcześniejszego hominina kopalnego, gdybyśmy go znaleźli? Jak odróżnimy go od najwcześniejszych kopalnych szympansów?
• Dlaczego rozwinęła się dwunożność?
• W jakim stopniu jedzenie mięsa ukształtowało ludzką ewolucję i umożliwiło zwiększenie rozmiaru mózgu?
• Kiedy ewoluował język?
• Dlaczego niektóre cechy, takie jak ciemno zabarwiona skóra, są bardziej rozpowszechnione w niektórych populacjach ludzkich niż w innych?
• Czy dobór naturalny nadal wpływa na populacje ludzkie? Czy ludzie wciąż się rozwijają?
Początki Człowieków (7)
DUBOIS I POZA
Pod koniec XIX i na początku XX wieku było wielu naukowców, którzy uważali, że kolebka ludzkości znajduje się w Afryce. Ale niektórzy, jak Ernest Haeckel i Eugène Dubois, dostrzegli podobieństwa między ludźmi a orangutanami z Indonezji i przewidywali, że tam właśnie miało miejsce ludzkie pochodzenie. Eugène Dubois był niezwykłą postacią w historii nauki o początkach człowieka. Spakował swoją rodzinę i swoje życie i przeniósł się do Holenderskich Indii Wschodnich, kierując się, jak wydawało się jego kolegom, fanatycznym przeczuciem, że odkryje tam "brakujące ogniwo". Ale w 1891 roku udowodnił, że wszyscy się mylili, odkrywając skamieniałości homininów na wyspie Jawa. Po trzech latach analiz nazwał swoje znalezisko Pitekantropus, co łączy w sobie greckie określenie małpy (pithekos) i człowieka (anthropos). Jego znalezisko zostało później przemianowane na Homo erectus, gatunek, który miał ciało podobne do człowieka, ale mniejszy mózg przypominający małpę. Wiele kolejnych ekspedycji rozpoczętych w latach dwudziestych XX wieku odzyskało znacznie więcej szczątków H. erectus z innych miejsc w Indonezji, a także ze słynnego stanowiska "człowieka pekińskiego" w Zhoukoudian w Chinach, gdzie wyhodowano ponad trzydzieści osobników o masie od 500 do 250 kya. O wiele bardziej podobnych do małp człekokształtnych przodków niż skamieniałości Dubois nie słyszano aż do 1924 r., kiedy Raymond Dart przeanalizował małą czaszkę ze skamieniałym mózgiem, znaną jako "Dziecko z Taung" z jaskini Sterkfontein w RPA. Okaz był bardziej prymitywny niż H. erectus i wspierał hipotezę pochodzenia afrykańskiego. Szczątki nazwał Australopithecus africanus, co oznacza "południowy małpolud z Afryki". Krytycy, a także zwolennicy miejsca Człowieka z Piltdown w ewolucji człowieka twierdzili, że czaszka była zbyt podobna do małpy, aby należeć do ludzkiego rodu. Ale Dart wykazał, że chociaż nie posiadał cech charakterystycznych, które mógłby zaoferować dorosły, dziecko z Taung miało ludzkie zęby i czaszkę, które różniły się od małp człekokształtnych. W 1959 roku Robert Broom rozpoznał już solidne skamieliny australopita (dziś nazywanego rodzajem "Paranthropus") z południowoafrykańskich stanowisk jaskiniowych, ale prawie kompletna czaszka znaleziona przez Louisa i Mary Leakey w 1959 roku zrobiła znacznie większy plusk. Odkrycie przez Leakeya OH 5 z wąwozu Olduvai w Tanzanii wykazało niewielkie oznaki deformacji i miało kompletną twarz. Natychmiast otrzymał przydomek "Człowiek z dziadka do orzechów", a to znalezisko, po prawie dwudziestu latach poszukiwań ludzkich skamieniałości, doprowadziło do kaskady odkryć więcej skamieniałości homininów i narzędzi kamiennych z wąwozu Olduvai, w tym wielu okazów wczesnego Homo. Wkład Louisa Leakeya (1903-1972) i Mary Leakey (1913-1996) w badania nad ewolucją człowieka oznacza początek współczesnej epoki paleoantropologii, ponieważ zanurzyli się oni w Afryce jako odkrywcy i odkrywcy, oddani odkryciu pochodzenia człowieka. Znaleźli wiele miejsc. Zainspirowali i dosłownie rozpoczęli liczne kariery innych naukowców w poszukiwaniu ludzkich początków i ewolucji. Byli znakomici w nagłaśnianiu swoich niezwykłych znalezisk. W swoje wyprawy i badania włączyli miejscową ludność. A co najważniejsze, utrwalili Afrykę Wschodnią jako kolebkę ludzkości. Richard Leakey i Meave Leakey przejęli kadencję Louisa i Mary w Kenii i nadal znajdują ekscytujące skamieliny. Po założeniu że Kenia i Tanzania są skarbnicami miejsc homininów, paleontolodzy zaczęli odkrywać bogactwo materiału również w Etiopii. Rok 1974 to rok odkrycia przez Donalda Johansona szkieletu Australopithecus afarensis zwanego "Lucy", który był najbardziej kompletnym wczesnym homininem znanym wówczas nauce. W Kenii, w latach 70., 80. i 90., szczególnie odnosząca sukcesy grupa paleontologów, nazywana "Gangiem Hominidów", dostarczała stały strumień skamieniałości hominidów. Zespół kierowany przez Richarda Leakeya (syna Louisa Leakeya), Alana Walkera, Meave Leakey, Johna Harrisa i Kamoya Kimeu zebrał liczne czaszki i kości pozaczaszkowe Homo erectus, Homo rudolfensis, Homo habilis, Paranthropus boisei, Paranthropus aethiopicus. W 1985 roku natrafili na najbardziej kompletny szkielet wczesnego hominina, jaki kiedykolwiek znaleziono, chłopca z H. erectus, zwanego "chłopcem Nariokotome" lub "chłopiec z Turkana", nazwanym tak od miejsca i brzegu jeziora, w którym został odkryty.
Brakujące ogniwo
Zawsze będą brakujące ogniwa lub luki w zapisie kopalnym, w których spodziewamy się znaleźć formy przejściowe. Organizmy, od grzybów, przez ryby, po rybaków, wyewoluowały przystosowania do przetwarzania zwłok w energię, zapobiegając skamienieniu większości organizmów. Pojęcie "brakujących ogniw" jest utrwalane, ponieważ za każdym razem, gdy odkrycie skamieniałości wypełnia lukę w zapisie kopalnym, tworzone są dwie kolejne luki. Następnie, gdy zostaną znalezione skamieliny, które wypełnią te luki, powstają kolejne cztery i tak dalej. Przeciwnicy ewolucji wykorzystują to pozornie niekończące się poszukiwanie brakujących ogniw, aby argumentować przeciwko wiarygodności ewolucji i produktywności paleontologii. To nieporozumienie najlepiej chyba nazwać "związkami mitów", ponieważ rzeczywiście istnieją niezliczone formy przejściowe, od ptaków z zębami i kostnymi ogonami (Archaeopteryx), przez wieloryby z małymi nogami (Ambulocetus), po małpy chodzące prosto (Australopithecus). Terminy "brakujące ogniwo" i "forma przejściowa" również są bardzo mylące. Sugerują, że organizm nie był dobrze przystosowany i po prostu czekał, a nawet dążył do ostatecznej ewolucji w nowoczesną formę. Są one również błędnie interpretowane jako oznaczające, że formy przodków są w jakiś sposób równymi mieszankami lub mieszankami ich współczesnych potomków.
Początki Człowieków (6)
PILTDOWN
Wcześni badacze ewolucji, w tym Darwin, przewidywali, że skamieliny naszych najwcześniejszych przodków będą podobne do małp. Jednak zanim zapis kopalny był tak bogaty jak dzisiaj, powszechnie zakładano, że homininy wyewoluowały duże mózgi, zanim wyewoluowały ciała podobne do ludzi. Wyglądało to tak, jakby homininy postanowiły swoim bystrym intelektem chodzić w pozycji wyprostowanej (tak jakby "w górę" było "właściwe") i używać narzędzi z uwolnionymi rękami. Ze względu na istniejące obecnie dowody wiemy, że dwunożność pojawiła się po raz pierwszy około 6 milionów lat temu u hominidów z małymi mózgami, które nie wytwarzały narzędzi kamiennych, a znaczna ekspansja mózgu homininów nastąpiła dopiero po 2 milionach lat. Ta wczesna hipoteza o wielkości mózgu i jego żarliwym podążaniu, połączona z chęcią uwierzenia, że pierwsi inteligentni ludzie pochodzili z Wielkiej Brytanii, uczyniła osławiony okaz skamieniałości znany jako "człowiek z Piltdown" tak potężnym oszustwem. W latach 1908-1912 w pobliżu wioski Piltdown w Sussex w południowej Anglii dokonano szeregu odkryć paleontologicznych i archeologicznych. Wśród szczątków dawno wymarłych ssaków, takich jak hipopotamy i nosorożce, był podobny do człowieka kawałek kości czaszki i przypominający małpę fragment szczęki, który Charles Dawson (amatorski antykwariusz i paleontolog o znacznej reputacji) nazwał "Eoanthropus dawsoni". Wielu uczonych z radością zaakceptowało Człowieka z Piltdown (tak nazywano go), ponieważ pokazało, że duże mózgi wyewoluowały wcześnie jako kamień węgielny ewolucji człowieka, a ludzie z dużymi mózgami wyewoluowali najpierw w Anglii. Nigdy nie było pełnej naukowej zgody co do tego, jak dopasować Człowieka z Piltdown do drzewa genealogicznego homininów lub jak zrekonstruować anatomię, ponieważ brakowało wówczas materiału porównawczego. Nawet gdy dodatkowe skamieniałości homininów zostały odkryte w Chinach i Afryce w latach 20. i 30. XX wieku, brakowało im zachowania tych samych części dla porównania, a ponadto coraz więcej skamieniałości homininów wykazało, że ludzkie zęby wyewoluowały wcześnie, podczas gdy ludzki mózg ewoluował znacznie później. Wraz z nagromadzeniem dowodów gdzie indziej anomalia z Piltdown stała się frustrująca i wprawiająca w zakłopotanie dla nauki. Wraz z nadejściem i zastosowaniem nowych technologii w latach 40. problem Piltdown zaczął odsłaniać się jak tajemnica. Analiza fluoru wykazała, że kość czaszki i szczęka nie pochodziły od tego samego osobnika. Po śmierci zwierzęcia azot w jego kościach zostaje zastąpiony fluorem, który jest wchłaniany przez wody gruntowe. Poziomy fluoru w obu kościach byłyby takie same, gdyby pochodziły od tego samego zwierzęcia, ale były różne. Co więcej, analizy datowania uranu wykazały, że kości były młodsze niż 50 000 lat, co jest o wiele za niedawne dla człowieka-małpy. W 1953 roku Joseph Weiner, Kenneth Oakley i Wilfred Le Gros Clark opublikowali "Rozwiązanie problemu Piltdown" w Biuletynie Brytyjskiego Muzeum Historii Naturalnej. Ich analiza wykazała, że szczęka z Piltdown pochodziła od młodej samicy orangutana, a czaszka należała do współczesnego człowieka. Piltdown było niczym więcej jak podróbką stworzoną tak, by wyglądała na starożytną, dzięki artystycznemu zastosowaniu chemikaliów do barwienia. Analiza mikroskopowa wykazała, że zęby zostały opiłowane, aby wyglądały na bardziej niejednoznaczne i ludzkie. Część szczęki, która łączy się z czaszką, została oderwana, aby ukryć, że nie mogą do siebie pasować. Chociaż mistyfikacja taka jak Człowiek z Piltdown nie mogła się wydarzyć w dzisiejszych czasach, sprawca fałszerstwa z Piltdown jest wciąż nieznany. Charles Dawson jest prawdopodobnie najlepszym przypuszczeniem, ponieważ był odpowiedzialny za różne odkrycia w Piltdown i był również związany z przypadkami oszustw związanych z artefaktami w innych witrynach. Jednak lista podejrzanych wciąż rośnie i obejmuje autora Sir Arthura Conan Doyle′a, który brał udział w wykopaliskach w Piltdown, a także Martina Hintona, byłego kustosza skamieniałości w British Museum (Historii Naturalnej). Niedawno odkryte resztki materiałów laboratoryjnych świadczą o zaangażowaniu Hintona w procesy geologiczne, które niszczą i plami skamieliny, a on mógł być wspólnikiem Dawsona w fałszerstwie.
Początki Człowieków (5)
Przodkowie
Poza epicką rolą Matki Natury nie ma znanych pramatek, które bezpośrednio przyczyniły się do powstania człowieka i nauki o ewolucji przed XX wiekiem. Co więcej, znaczna część teoretycznego i filozoficznego tła nauki o pochodzeniu i ewolucji człowieka ma swoje źródło w Europie, ponieważ tam narodziła się nauka, jaką znamy. Ale zarówno dla Europejczyków, jak i dla nie-Europejczyków, poszukiwanie początków człowieka i zrozumienia ewolucji człowieka jest głęboko słyszalne dla każdego, kto kiedykolwiek zastanawiał się nad ludzką naturą i jak ludzie powstali. Skamieniałości wzbudzają naszą ciekawość, a pisemne zapiski, folklor i kolekcja artefaktów wskazują, że ludzie zawsze fascynowali się rzeczami, które wykruszają się z ziemi. Chińscy uzdrowiciele nadal kierują się starożytnym przekonaniem, że "smocze kości" - skamieliny dinozaurów i innych zwierząt - są skutecznymi lekami. Dekoracje na ceramice i zapiski pisane ujawniają, że w przypadku klasycznych Greków i Rzymian skamieniałe kości z ziemi wywoływały pytania i pomagały wspierać przekonania o ich własnym pochodzeniu. Wybitne rodziny wywodziły swoje rodowody od bohaterów o znaczeniu mitologicznym. Skamieliny dużych wymarłych zwierząt, takich jak starożytne słonie, z miocenu i plejstocenu potwierdziły ich przekonanie, że poprzedzali je olbrzymy i przerażające bohaterowie mitów. Dziś muzea na całym świecie prezentują skamieniałości z ich regionu i chwalą się, że ich część świata ma kluczowe znaczenie dla pochodzenia człowieka. Ta starożytna praktyka została wymyślona przez cesarza Augusta (63 r. p.n.e - 14 r. n.e.), który założył pierwsze na świecie muzeum paleontologiczne. Zawierała kości olbrzymów i broń starożytnych bohaterów, a wszystko to wykorzystywane jako propaganda wzmacniająca jego rodzące się imperium rzymskie. Ale zanim pierwsi paleontolodzy i ich muzea, Platon i Arystoteles (VI i V wiek p.n.e.) ustanowili pierwszą zapisaną filozofię dotyczącą ludzkiej natury i ludzkiego pochodzenia. Uważali ludzi za część naturalnego świata, tak jak inne organizmy, i umieścili ich na szczycie "Wielkiego Łańcucha Bytu". Wraz z rozprzestrzenianiem się chrześcijaństwa łańcuch został zaadoptowany jako drabina z Bogiem na szczycie, aniołami tuż pod nim, ludźmi podzielonymi na szereg kategorii rasowych poniżej aniołów, naczelnymi nieludzkimi pod nimi, ssakami, następnie gadami, płazami i rybami, z roślinami zakorzenione na dole, na nieożywionych przedmiotach, takich jak skały. Teologia chrześcijańska zdominowała średniowiecze Europy Zachodniej, a świat był postrzegany jako wytwór planu Bożego. Gatunki były uważane za stałe i niezmienne i wyglądały dzisiaj tak, jak zawsze. Uważano, że ludzie są oddzieleni od otaczającego ich naturalnego świata i zostali stworzeni przez Boga w ich obecnej formie z językiem i kulturą. Zakładano, że biblijne stworzenie miało miejsce bardzo niedawno z powodu obliczeń uczonych, takich jak biskup Ussher. Podsumował wieki pokoleń ludzi w Starym Testamencie i obliczył, że stworzenie świata opisane w Księdze Rodzaju nastąpiło w 4004 r. p.n.e. Rewolucję naukową XVII wieku zapoczątkowali Kopernik i Galileusz, którzy zdali sobie sprawę, że świat kręci się wokół słońca, a nie na odwrót. Porzucono również długo utrzymywany pogląd, że Ziemia jest płaska. Co więcej, odkrywcy coraz częściej wchodzili w kontakt z ludźmi za oceanami i poznawali ich unikalne tradycje kulturowe i niezachodnie sposoby bycia ludźmi. Sztywna biblijna interpretacja historii naturalnej zaczęła sprawiać problemy naukowcom zainteresowanym wyjaśnianiem otaczającego ich świata. Carl von Linné, lepiej znany pod zlatynizowanym imieniem Carolus Linnaeus (1707-1778), zrobił wielki krok w kierunku współczesnego stanu człowieka. Początki nauki, kiedy nadał ludziom nazwę rodzajową i gatunkową, tak jak dał gady i dęby. Linneusz stworzył dwumianową metodę klasyfikacji organizmów i nazwał ludzi Homo (człowiek) sapiens (mądry). Linnejski system taksonomii jest dziś uniwersalny. Georges Cuvier (1769-1832) był pierwszym, który zastosował zasady naukowe (ustanowione przez Francisa Bacona w 1620) do paleontologii, a konkretnie do paleontologii kręgowców. Zdał sobie sprawę, że katastrofy, takie jak te opisane w Starym Testamencie Biblii, były prawdopodobnie odpowiedzialne za wyginięcie wielu gatunków znalezionych jako skamieniałości. W teorii "katastrofizmu" Cuviera obszary, które doświadczają masowego wymierania, są uzupełniane nowymi formami migrującymi z sąsiednich regionów. Jednak Cuvier nigdy nie wyobrażał sobie powolnych zmian w głębokim czasie i właśnie dlatego wkład Charlesa Lyella (1797-1875) w "uniformitarianizm" uczynił go założycielem współczesnej geologii. Teoria Lyella, która obowiązuje dzisiaj, głosiła, że te same fundamentalne procesy naturalne, które zachodzą dzisiaj (zarówno katastrofalne, jak i powolne, stopniowe zmiany) zawsze miały miejsce w przeszłości i nadal będą zachodzić w przyszłości. Tak więc dzisiejsza Ziemia wygląda znacząco odmiennie od tego, jak wyglądała miliony lat temu z powodu procesów takich jak erozja i wulkanizm, które kształtują góry i wąwozy. Lyell był jednym z pierwszych, którzy argumentowali, że Ziemia jest rzeczywiście bardzo stara i nie może mieć zaledwie 6000 lat, jak obliczył biskup Ussher. Darwin przeczytał Zasady geologii Lyella (1830) podczas swojej historycznej podróży na H.M.S. Beagle i nacisk Lyella na starożytny wiek ziemi, głęboki czas i stopniowy, powolny charakter procesów geologicznych miał na niego duży wpływ. Zanim Karol Darwin sformułował swoją teorię ewolucji przez dobór naturalny w 1859 roku w O powstawaniu gatunków, skamieniałości neandertalczyków i kromaniończyków wywołały już gigantyczne fale w społeczności naukowej. W 1829 r. w Engis w Belgii odkryto pierwsze szczątki neandertalczyka, ale dopiero później rozpoznano czaszkę dziecka. W 1848 roku w kamieniołomie Forbesa niedaleko Gibraltaru odkryto dorosłą czaszkę neandertalczyka, która również nie została wówczas rozpoznana. W 1856 r. w kamieniołomie Feldhofer w dolinie Neander w Niemczech odkryto cały szkielet neandertalczyka. Neandertalczyk jako oddzielna, nieludzka istota - w większości oddzielona różnicami w kształcie twarzy i czaszki, a także wytrzymałością i solidnością szkieletu - została założona w 1864 roku i nazwana zgodnie ze swoim rodzinnym regionem: Homo neanderthalensis. Skamieliny współczesnych ludzi nie były nawet znane, zanim neandertalczycy zostali osiedleni jako prehistoryczni "jaskiniowcy". W końcu w 1868 r. w schronie skalnym z Cro-Magnon w Les Eyzies w Dordogne we Francji odkryto szkielety skamieniałych współczesnych ludzi, stąd przydomek "Cro-Magnon" oznaczający epokę lodowcową lub późnoplejstoceńskich współczesnych ludzi z Europy. Pierwsi Cro-Magnonowie znani nauce byli niewiele młodsi od neandertalczyków, ale mieli wyraźnie ludzkie cechy i nie posiadali żadnych prymitywnych cech widocznych na czaszce i szkielecie neandertalczyka. W przeciwieństwie do starożytnych Greków, którzy często mylili się, interpretując skamieniałości nieludzkie jako swoich przodków, wielu z pierwszych naukowców, którzy interpretowali najwcześniejsze szczątki neandertalczyka i Homo erectus znalezione w połowie XIX wieku, odmówiło uznania ich roli w ewolucja człowieka. Tak zwane logiczne wyjaśnienia zwykle oferowane dla dziwnie ukształtowanych skamieniałości były takie, że były to chore szczątki współczesnego człowieka lub ranne szczątki żołnierzy, a nie starożytne lub wymarłe humanoidy. I odwrotnie, naukowcy, którzy zaakceptowali dowody paleontologiczne na ewolucję człowieka, stali się pierwszymi paleoantropologami. Co więcej, ci mężczyźni nie byli już przypadkowymi turystami dyscypliny. Oznacza to, że nie byli już ograniczeni do kontemplowania niesamowitych znalezisk dokonanych tuż za drzwiami ich szpitala lub wiejskiej chaty. Ci ludzie byli uczonymi w anatomii człowieka i postanowili znaleźć wyraźny cel odnalezienia starożytnych skamielin ludzkich lub z wyraźnym celem znalezienia dowodów na ewolucję człowieka. Na przykład Thomas Henry Huxley (1825-1895) napisał książkę zatytułowaną, która dzisiaj może być mylnie zinterpretowana jako podręcznik rekreacji na świeżym powietrzu, a nie epos ewolucyjny: Dowody na miejsce człowieka w przyrodzie (1863). Huxley był pierwszym naukowcem, który wyraźnie porównał anatomię ludzi i małp człekokształtnych, pracując pod tym, co stało się jego słynnym zarzutem:
Kwestia pytań dla ludzkości - problem, który leży u podstaw wszystkich innych i jest o wiele bardziej interesujący niż jakikolwiek inny - jest ustalenie miejsca, jakie człowiek zajmuje w przyrodzie i jego relacji do wszechświata rzeczy. (TH Huxley, 1863)
Huxley twierdził, że ludzie i afrykańskie małpy człekokształtne są bardzo blisko spokrewnione, a nawet bardziej niż z małpami azjatyckimi. Postawił zatem hipotezę, że ludzkie pochodzenie można znaleźć w Afryce - przepowiednię tę później poparł Darwin w swojej książce The Descent of Man (1871) i którą potwierdzili również przyszli paleoantropolodzy.
Początki Człowieków (4)
METODA NAUKOWA
Kiedy paleoantropolog znajduje skamieniałość, stosuje naukową metodę rozszyfrowania jej miejsca w historii ewolucji. Na podstawie wcześniejszej wiedzy uzyskanej z pracy innych naukowców oraz własnych obserwacji formułuje hipotezę. Następnie zbiera dowody lub dane, aby sprawdzić swoją hipotezę. Na tym etapie chemik przeprowadzałby eksperyment, ale paleoantropolog zbiera dowody eksperymentu ewolucji, analizując inne skamieniałości i kości żywych gatunków, które są podobne. Jeśli zebrane przez nią dane potwierdzają jej hipotezę, paleoantropolog i inni badacze powtarzają proces naukowy, aby ustalić, czy dodatkowe dowody mogą wspierać tę samą hipotezę. Jeśli tak i jeśli hipoteza nadal jest wspierana, to ta hipoteza zamienia się w teorię. Im silniejsze jest poparcie dla teorii, tym silniejsze muszą być dowody przeciwko niej, aby ją sfalsyfikować. W miarę zdobywania poparcia w postaci powtarzanych testów, teoria ta w końcu stanie się tak silna, że stanie się częścią wcześniejszej wiedzy, którą inni naukowcy stosują do formułowania nowych hipotez. Cykl ten trwa w nieskończoność, gdy hipotezy są testowane i albo wspierane, albo nie, teorie są obalane przez odkrycie dowodów, które je obalają, wraz z zupełnie nowymi hipotezami rodzącymi się, gdy teorie się kruszą, a niektóre teorie wytrzymują próbę czasu. Przed zebraniem jakichkolwiek skamieniałości paleoantropolog wybiera stanowisko terenowe w oparciu o typy skamieniałości, które może wykorzystać do sprawdzenia swoich hipotez. Tak więc, jeśli jej hipotezy koncentrują się na podziale między rodowodami szympansów i ludzi, bada miejsce o znanym wieku geologicznym lub takim, który odpowiada wiekiem innym miejscom, które już wyprodukowały bardzo wczesne skamieniałości homininów. Szuka skał, które pasują do wieku genetycznego szacowanego czasu dla rozłamu około 6 milionów lat temu. Wraz z wszelkimi skamieniałościami szympansów lub przodków człowieka, paleoantropolog musi również obserwować i gromadzić inne rodzaje skamieniałości na tym stanowisku. Zachowane szczątki roślinne i zwierzęce pomogą w odtworzeniu dawnego środowiska. Charakter warstw skalnych wskaże również, czy osady zostały zdeponowane przez jezioro, deltę, rzekę, wąwóz, norę zwierzęcą, jaskinię lub wiele innych scenariuszy pochówku. (Do niedawna homininy nie były znajdowane wewnątrz celowo wykopanych grobów.) Tego typu rekonstrukcje środowiskowe umieszczą gatunek w kontekście środowiskowym i dostarczą również dodatkowych informacji o ewolucji małp i ludzi, których skamielina nie może dostarczyć sama. W laboratorium paleoantropolog przeanalizuje anatomię skamieniałości, rejestrując pomiary (np. długość i szerokość kości i zębów) oraz zaglądając do wnętrza kości za pomocą technologii obrazowania, takiej jak rentgenowska tomografia komputerowa (tj. "CT" lub skanowanie "cat"). Dane te są następnie porównywane z innymi skamieniałościami i szkieletami współczesnych małp człekokształtnych i ludzi. Paleoantropolog sprawdzi hipotezę, że zebrana przez nią skamielina jest bardziej spokrewniona z szympansami lub z ludźmi, sprawdzając, czy cechy anatomiczne są bardziej podobne do siebie. Jeśli cechy skamieniałości bardziej przypominają współczesne szympansy niż ludzkie, to zostanie ona umieszczona w ewolucyjnym rodowodzie szympansów. To umiejscowienie jest hipotezą, którą można obalić dodatkowymi dowodami. W końcu możliwe jest, że małpopodobna istota żyjąca 6 milionów lat temu nie była ani przodkiem szympansów, ani ludzi, ponieważ mogła należeć do linii, która wymarła i nie przyczyniła się do naszej ewolucji ani do ewolucji naszych małpich kuzynów. Nadrzędną ramą w poszukiwaniu początków człowieka jest teoria, która osiągnęła stan faktyczny: ewolucja przez dobór naturalny. Naukowo rygorystyczne badanie początków i ewolucji człowieka mogło rozkwitnąć po tym, jak Karol Darwin wniósł ten wkład, częściowo dlatego, że przedstawił hipotezy do sprawdzenia. Na przykład w The Descent of Man (1871) Darwin postulował, że skamieniałości ostatniego wspólnego przodka (LCA) wielkich małp człekokształtnych i ludzi można znaleźć w Afryce, ponieważ goryle, bonobo i szympansy, które są obecnie najbardziej ludzkimi zwierzętami, żyć tutaj. Zgodnie z aktualnym zapisem kopalnym, prognoza Darwina jest nadal słuszna i oczywiście może się okazać, że się myli, jeśli naukowcy znajdą dowody. Jest mało prawdopodobne, ale jednak możliwe, że jutro ktoś mógłby odkopać LCA w Azji i wywrócić wszystko do góry nogami. Taka jest podstawowa natura paleoantropologii i nauki.
Początki Człowieków (3)
NAUKA O LUDZKIM POCHODZENIU I EWOLUCJI
Niezależnie od tego, czy badają malowidła jaskiniowe z epoki lodowcowej, DNA szympansów, czy kości pierwszych maleńkich, przypominających wiewiórkę naczelnych sprzed 60 milionów lat, naukowcy zadają te same pytania: Skąd pochodzimy i jak się tu dostaliśmy? Chociaż poszukiwanie początków człowieka czerpie z badań wielu dyscyplin naukowych, ogranicza się głównie do antropologii. Szeroko definiowana jako "badanie ludzi", antropologia może obejmować każde dążenie naukowe, o ile koncentruje się na człowieku. Antropologia biologiczna (zwana również antropologią fizyczną) jest dziedziną antropologii, a wszystkie jej poddziedziny przyczyniają się bezpośrednio lub pośrednio do zrozumienia pochodzenia człowieka i ewolucji. Te poddziedziny obejmują badania naukowe genetyki, behawioru, biologii, ekologii i ewolucji ludzi i innych naczelnych, które podlegają dyscyplinom takim jak "paleoantropologia", która jest badaniem ewolucji człowieka i "prymatologia", która jest badaniem zachowania i ekologii żyjących naczelnych. Antropologia biologiczna nie jest osamotniona w poszukiwaniu początków człowieka. Inne dziedziny antropologii archeologii i antropologii kulturowej są również kluczowe dla zrozumienia pochodzenia i ewolucji człowieka. Ponieważ paleoantropologia rekonstruuje zamierzchłą przeszłość, jest nauką historyczną. Tak więc trudno jest wyciągnąć takie same wnioski, jakie chemicy lub biolodzy komórkowi mogą wyciągnąć z eksperymentów naocznych świadków. W paleoantropologii "Matka Natura" przeprowadziła eksperymenty, a tysiące lub miliony lat później naukowcy opisują starożytne eksperymenty niewidzialne i wyjaśniają wyniki na podstawie dowodów, które przetrwały.
Początki Człowieków (2)
Ułożenie układanki o początkach człowieka i ewolucji jest interdyscyplinarnym przedsięwzięciem. Dziedziny genetyki, paleontologii, paleoantropologii, archeologii, antropologii kulturowej, prymatologii, behawioru i ekologii zwierząt, anatomii, fizjologii, kinezjologii, psychologii, kognitywistyki, ekonomii i wielu innych oferują fragmenty układanki. Korzenie ludzkości sięgają początków wszechświata, ponieważ składamy się z atomów, zarania życia na ziemi, ponieważ jesteśmy na bazie węgla, pierwszej ryby, która zarabia na życie na lądzie, ponieważ jesteśmy czworonogami. małym ssakom, które przetrwały apokalipsę kończącą dinozaury, ponieważ karmimy się mlekiem jako niemowlęta i są futrzane (niektóre bardziej niż inne), małpom człekokształtnym, które z jakiegoś dziwnego powodu są dobre w kiwaniu się na tylnych łapach, i wreszcie doświadczonym dwunożnym z dużym mózgiem do opanowania tworzenia narzędzi i języka. Z pewnością nie można opisać w tym tomie całej historii wszechświata w odniesieniu do początków i ewolucji człowieka. Głównym celem studiów nad ewolucją człowieka (i głównym celem tutaj) jest z grubsza sześć milionów kluczowych lat od momentu, w którym nasza linia rodowodowa oddzieliła się od przodków, których dzielimy z szympansami, aż do pojawienia się pierwszych współczesnych ludzi w Afryce, tuż przed tym, jak nasze indywidualne i populacyjne historie zaczęły się rozchodzić. Chociaż większość terminów technicznych jest zdefiniowana w słowniku, są dwa, które wymagają szczególnej uwagi od samego początku. Homininy to zmodyfikowane małpy afrykańskie. Obejmują one żywych ludzi i wszystkich wymarłych potomków ostatniego wspólnego przodka z szympansami, które są w linii ludzkiej, w przeciwieństwie do linii szympansów. Skamieniałe homininy są albo naszymi bezpośrednimi przodkami, albo naszymi ewolucyjnymi kuzynami znajdującymi się na różnych gałęziach drzewa homininów od naszego. Większość homininów jest dwunożnych i ma stosunkowo duże mózgi, ale te najwcześniejsze mogły takie nie być, jeśli te cechy nie wyewoluowały natychmiast po oddzieleniu się od wspólnego przodka z szympansami. Ludzie są jedynym żyjącym gatunkiem homininów i różnią się od innych gatunków homininów naszą unikalną kombinacją złożonej kultury materialnej, zachowań społecznych, cech cielesnych i inteligencji. Tutaj będziemy odnosić się tylko do anatomicznie współczesnych ludzi lub Homo sapiens jako "ludzi". To są ludzie, których nazwalibyśmy "ludźmi", gdybyśmy stanęli oko w oko z jednym z nich. Innymi słowy, gdybyśmy cofnęli się wehikułem czasu do 200 000 lat temu, prawdopodobnie rozpoznalibyśmy wyprostowane istoty chodzące jako istoty ludzkie (nawet jeśli prawdopodobnie nie używały jeszcze języka ani symboliki tak jak my), ale nie ma zagwarantować, że połączylibyśmy się w ten sam sposób ze wszystkim, co żyło wcześniej, niekoniecznie nawet z neandertalczykami, którzy żyli obok niektórych populacji ludzkich do 30 000 lat temu. Chociaż niektórzy naukowcy nazywają wszystko w obrębie rodzaju ludzkiego (Homo) "ludźmi", rezerwujemy ten tytuł tylko dla siebie. Wydaje się logiczne, że jeśli ktoś ma badać pochodzenie człowieka, powinna istnieć powszechna zgoda co do tego, co dokładnie czyni człowieka "człowiekiem". Ale na pytanie "co definiuje ludzi lub co czyni ich wyjątkowymi?" wiele popularnych odpowiedzi zawiera się w sferze intelektualnej i emocjonalnej: miłość, śmiech, żal i strata, kreatywność, ciekawość, głód nauki, postrzeganie rzeczy niewidzialnych, pojęcie przyszłości, duchowość, samobójstwo i język. Nie są to typy cech, które można łatwo prześledzić w prehistorii za pomocą skamieniałości, artefaktów lub genetyki. Porównawcze obserwacje innych zwierząt pomagają nam zrozumieć naturę tych ludzkich cech, ale im więcej uczymy się, w szczególności od małp, tym bardziej odkrywamy, że bardzo niewiele z tego, co definiuje człowieka, jest wyłącznie człowiekiem. 2 procent naszego genomu, który czasami różni się od szympansów ,wydaje się zbyt mały, aby pomieścić wszystkie nasze różnice. Fizycznie mamy większy mózg z gęstymi włosami pokrywającymi czaszkę, a u samców również twarz. Nasze owłosienie na ciele jest drastycznie zredukowane, a naga skóra zawiera dużo więcej gruczołów potowych. Nasze ręce są o wiele bardziej zręczne, ale nasze ciała są znacznie słabsze niż szympansów. Mamy duże, rzucające się w oczy organy płciowe i chodzimy tylko na dwóch bardzo długich nogach, obutych sztywnymi, mocnymi stopami. Duża część białka naszych oczu, które jest bielsze niż u innych małp człekokształtnych, jest widoczna dzięki migdałowemu kształtowi naszych powiek. Behawioralnie polegamy na bardzo skomplikowanych wokalizacjach. Kontrolujemy ogień. Graffitujemy prawie wszystko, łącznie ze sobą. Ogólnie rzecz biorąc, pokojowo gromadzimy się w ogromnych grupach składających się głównie z niespokrewnionych osób. Wyzyskujemy i selektywnie rozmnażamy inne gatunki (a nawet członków własnych, w aranżowanych małżeństwach). Jesteśmy zręcznymi myśliwymi bez kłów i pazurów i potrafimy przystosować się do każdego środowiska na ziemi. Na kolejnych stronach śledzimy pochodzenie i ewolucję cech, które umożliwiły człowiekowi stanie się "człowiekiem". Jeśli te cechy zostaną kiedyś zmapowane w genomie, odkryjemy, że są one wyrażane lub regulowane przez mniej niż 2 procentową różnicę DNA, która oddziela nas od szympansów. W miarę jak dowiadujemy się więcej o naszym własnym genomie i genomach pokrewnych zwierząt, odkrywamy, że większość tego, co uważamy za unikalne dla naszego gatunku, prawdopodobnie pojawiło się niedawno, a ponieważ większość cech jest złożona i wyrażana przez wiele genów, unikalne cechy ludzkie będą wyzwanie do określenia na poziomie molekularnym, nie mówiąc już o prześledzeniu przez ewolucję. W żadnym wypadku nie można zmieścić całej dziedziny ludzkich początków w jednej księdze, więc podstawy i najważniejsze elementy materiału dowodowego są tutaj zawarte. Słownictwo anatomiczne, a także epoki, okresy i epoki czasu geologicznego można znaleźć w Dodatkach A i B. W tekście "Mya" i "Kya" będą skrótem określającym miliony lat temu i tysiące lat temu. Rozpoczynamy krótką historią poszukiwań początków człowieka i badań nad ludzką ewolucją. W wyniku samoświadomości ludzie prawdopodobnie byli ciekawi swojego pochodzenia, odkąd pierwszy hominid rozwinął zdolność umysłową do robienia tego. Jednak współczesne badania nad pochodzeniem człowieka nie rozpoczęły się poważnie aż do połowy XIX wieku, kiedy w Europie znaleziono pierwszych neandertalczyków. Na początku historii tej dziedziny kontrowersje wokół nowych odkryć były spowodowane niepewnością, czy ludzie w ogóle ewoluowali. Teraz, gdy nauka pokonała stare kontrowersje, kontrowersje wokół nowych odkryć odnoszą się jedynie do szczegółów związanych z ich interpretacją. O ile skamieniałości nie mają znaczników identyfikacyjnych, zawsze będą kontrowersje dotyczące ich interpretacji. Jednak w większości przypadków większość naukowców zgadza się na interpretację, która staje się paradygmatem roboczym, chyba że nowe odkrycie wywróci wszystko do góry nogami. Ta kolegialna debata nad nowymi znaleziskami jest niestety wciąż przerabiana, a następnie zachwalana przez antyewolucjonistów jako dowód na istnienie błędów w teorii ewolucji. Ponieważ koncentrujemy się na procesach ewolucji, przedstawiamy zarys teorii ewolucji i omawiamy sformułowanie Darwina teorii ewolucji poprzez dobór naturalny, a także dobór płciowy. Koncentrujemy się na tym, jak dziś rozumiemy ewolucję z naszym obecnym zrozumieniem genetyki. W tym rozdziale przyjrzymy się dowodom, które zebraliśmy z otaczającego nas żyjącego świata na pochodzenie i ewolucję człowieka, więc omówione zostaną metody klasyfikacji i tworzenia drzew genealogicznych, a następnie omawiane są grupy w obrębie Naczelnych Zakonu, których jesteśmy członkiem. Bez podróży w czasie nigdy nie można być naprawdę pewnym, jak wyglądało życie, zanim po raz pierwszy zarejestrowano ruchome obrazy lub zanim ludzie zaczęli spisywać historię. Jednak stare kości i kamienie dostarczają zdumiewającej ilości informacji o pochodzeniu i ewolucji człowieka. Poźniej dokopujemy się do nagich kości naszych poszukiwań dowodów na ludzkie pochodzenie, do skamielin i zapisów archeologicznych. Spotykamy naszych przodków i bliskich kuzynów i sortujemy pozostawione przez nich śmieci. Omawiamy skamieniałości i artefakty, które przypadkowo zostały zachowane, które ciężką pracą i odrobiną szczęścia zostały odkryte i które dzięki starannej analizie naukowej zostały zinterpretowane jako dowód na naturę naszego pochodzenia i ewolucji. Architektura i mechanika naszego ciała to tylko część bardzo niedocenianej wiedzy, którą badacze codziennie gromadzą na temat naszej ewolucji i miejsca w naturze. Ustalenie metod geologicznych na początku rozdziału pozwala spojrzeć na skamieniałości z odpowiedniej perspektywy. Wczesne skamieniałe naczelne, małpy i małpy człekokształtne są krótko zwiedzane, a następnie po dotarciu do homininów są one podzielone na podpozycje według gatunków. Omawiane są tylko ogólne cechy anatomiczne, ale należy pamiętać, że naukowcy opierają rozróżnienia gatunkowe i interpretacje anatomiczne na dokładnych pomiarach i analizach statystycznych. Ponieważ tak wiele teorii ewolucyjnych opiera się na współczesnej genetyce, pokażemy postępy, jakie poczyniła genetyka w naszym zrozumieniu pochodzenia człowieka i ewolucji. Dzięki DNA można prześledzić pochodzenie człowieka aż do początków życia i organizmów wielokomórkowych. Wraz z coraz silniejszą biotechnologią, poszukiwanie pochodzenia człowieka nie opiera się już tylko na zakurzonych skamielinach i artefaktach. Dziś artefakty w genomie są tak samo ważne jak te zakopane w ziemi. W epoce, w której precyzja i zakres analizy molekularnej stale wzrasta, możemy wykorzystać ją jako narzędzie do śledzenia naszej ewolucji. Wiedza o genach i dziedziczeniu jest wykorzystywana do zbadania zastosowania zegarów molekularnych do określenia czasów podziału linii w prehistorii, roli DNA w określaniu naszego afrykańskiego pochodzenia, a także ewolucji niektórych niedawnych adaptacji ludzkich. Chociaż starożytne DNA nie jest technicznie nowoczesnym dowodem, techniki, za pomocą których jest ono ekstrahowane i analizowane, są nowoczesne, dlatego zostało tutaj uwzględnione. Syntetyzujemy dowody z żywych modeli zwierzęcych, skamieniałości, artefaktów i genetyki, aby prześledzić pochodzenie i ewolucję niektórych podstawowych cech człowieka. Rozważ ten rozdział jako plan budowy człowieka od małpiego przodka, w przeciwieństwie do budowania szympans z tego samego przodka. Ewolucja wielu, jeśli nie wszystkich, cech człowieka jest w dużym stopniu zależna od ewolucji innych. Na przykład, przyjęcie nawykowego dwunożności pozwoliło doborowi działać na anatomię nowo uwolnionych rąk, które stały się jeszcze bardziej biegłe w używaniu i wytwarzaniu coraz bardziej skomplikowanych narzędzi. Dlatego nie można adekwatnie badać dwunożności bez zrozumienia archeologicznego zapisu narzędzi kamiennych. Nasi przodkowie zostali ukształtowani przez te same naturalne procesy i siły ewolucyjne, co wszystkie inne organizmy artystyczne. Ale pomimo rozproszenia na całym świecie, złożoności kulturowej, innowacyjnych modyfikacji, które wprowadzamy w naszym otoczeniu i wielkiego wpływu, jaki wywieramy na środowisko, czy te same siły działają obecnie na ludzi? jak i kiedy nastąpiła kolonizacja świata? Czy będziemy mieli czas na znacznie większą ewolucję, czy wyginiemy, zanim się dowiemy? A gdyby mieli szansę zrobić to jeszcze raz, czy ludzie nadal by ewoluowali? Badania ewolucyjne są rewolucyjne. Dzięki dodatkowym odkryciom z wykopalisk i wykorzystaniu nowych technologii, hipotezy dotyczące pochodzenia i ewolucji człowieka można nie tylko testować i ponownie testować z wielu różnych perspektyw naukowych, ale są one stale ulepszane, aby odzwierciedlić napływ nowych dowodów. Jutro ktoś może znaleźć nowy element układanki, który albo wzmocni to, co większość ludzi już wiedziała, albo całkowicie wywróci ich perspektywę i zmusi do przemyślenia całej sprawy. Nowy gatunek skamieniałości lub przełom genetyczny mogą dosłownie z dnia na dzień zmienić sposób, w jaki myślimy o naszym pochodzeniu i ewolucji. Jeśli, hipotetycznie, ktoś jutro odkryje skamieniałego hominina z 9 milionów lat temu, który chodził wyprostowany, to historia będzie musiała zostać napisana od nowa (ponieważ w obecnej formie przytłaczające dowody wskazują na pochodzenie hominina około 6 milionów lat temu). Ale bez względu na przyszłość nauki o ludzkim pochodzeniu i ewolucji, informacje zawarte w tym tomie służą jako podstawa do zrozumienia, dlaczego takie odkrycie byłoby znaczące i jak wpłynęłoby na obecne zrozumienie oparte na przytłaczających dowodach ze skamieniałości, zapisy archeologiczne i genetyczne.
Początki Człowieków
Krótka odpowiedź na pytanie o pochodzenie człowieka została już poddana triangulacji przez genetykę, paleontologię i archeologię: 200 000 lat temu w Afryce Subsaharyjskiej. Dlaczego więc potrzebujemy tekstu tylko po to, by wprowadzić temat? Ponieważ długa odpowiedź to wciąż wydłużająca się saga, która zawiera tyle rozdziałów, ile jest cech składających się na gatunek ludzki. Różne aspekty Homo sapiens powstały w różnym czasie, aby zbudować stworzenia, którymi jesteśmy dzisiaj. Trzy maleńkie kości w naszych uszach, które służą do słyszenia, wyewoluowały ponad 100 milionów lat temu, ale nie zaczęliśmy produkować instrumentów muzycznych przed co najmniej 100 000 lat temu. Każde nabycie cechy ludzkiej wpływało na wydarzenia znajdujące się na dalszej ścieżce ewolucji człowieka. To z powodu tego szerszego poszukiwania początków człowieka jest coś więcej do omówienia niż tylko "200 000 lat temu w Afryce subsaharyjskiej". Gdyby każdy człowiek narysował swoje drzewo genealogiczne aż do początków życia na planecie Ziemia, historia każdego człowieka byłaby identyczna od Wielkiego Wybuchu do niedawna, kiedy nasze indywidualne historie - gałązki naszych plemiennych rodów - oddzieliły się od gałęzi innych grup ludzkich i stały się odrębne geograficznie i genetycznie. Tak jak człowiek może prześledzić swoje kręcone włosy z pokolenia na pokolenie, patrząc na zdjęcia swoich kuzynów, dziadków i pradziadków, gatunek ludzki może prześledzić swoje korzenie, patrząc na skamieniałości i geny, które ujawniają naszych wspólnych przodków z naszym wymarłym homininem przodków i naszych "kuzynów" - małpy człekokształtne, małpy, myszy, konie, ryby, robaki, kukurydza, śluzowce, bakterie i wszystkie żywe stworzenia, które ludzie kiedykolwiek chcieli nazwać. To z powodu naszego intelektu często bardzo łatwo zapominamy, że jesteśmy członkami królestwa zwierząt, działającymi w ramach tych samych podstawowych procesów ewolucyjnych i kształtowanymi przez te same, co wszystkie inne żywe istoty. Pomimo długiej listy różnic, od których wyobrażamy sobie, że nas oddzielają lub wywyższają od innych zwierząt, różnimy się od szympansów o mniej niż 2 procent naszego kodu genetycznego (co nie jest dużo, biorąc pod uwagę, że ma on długość 3 miliardów par zasad). Z pewnością to nasze różnice w stosunku do innych zwierząt definiują nas jako ludzi, ale to nasze fundamentalne podobieństwa do tych zwierząt ostatecznie pozwoliły nam stać się ludźmi. Wiedząc, że jesteśmy prawi, gadające małpy afrykańskie nie muszą wpędzać nas w kryzys egzystencjalny, nie ze świadomością, że każdy z nas jest wyjątkowy. Żadne dwie osoby nie mają dokładnie tego samego DNA, nawet dorosłe bliźnięta. Nikt w przeszłości ani w przyszłości nigdy nie będzie miał tego samego kodu genetycznego, co jakakolwiek inna osoba w przeszłości lub przyszłości, ani żaden inny organizm. Każdy z nas jest wynikiem jednego udanego, nieprzerwanego łańcucha życia, który rozpoczął się 3 miliardy lat temu. Zamiast wykorzystywać tę dziwnie bystrą inteligencję, by tarzać się w naszych zwykłych naczelnych, powinniśmy zamiast tego podziwiać, co dobór naturalny i inne siły biologiczne wytworzyły u przodka z mózgiem małpim. Jako prawdopodobnie jedyne samoświadome stworzenia na planecie, szokujące jest to, jak mało wiemy o sobie. Wiele osób zapytanych, powie, że są "homo sapien", błędnie zakładając, że gatunek w naszej naukowej nazwie to liczba mnoga. Niektórzy ludzie potrafili zmontować części samochodu sportowego aż do ostatniego łożyska kulkowego, ale nie mogli zlokalizować własnych kanałów półkolistych, nie mówiąc już o opisaniu tego, co robią. (Są w uszach, ale nie mają nic wspólnego ze słyszeniem. Są zaangażowane w wyraźne widzenie, podczas gdy głowa porusza się.) Poprzez ustne i pisemne historie unieśmiertelniamy nasze rodzinne genealogie, nawet nazywamy nasze dzieci po ich przodkach, ale rzadko przeszkadza nam zapamiętanie bardzo długich, dziwnie wymawianych imion naszych wymarłych ewolucyjnych przodków. Obok książek do anatomii na półkach w naszej bibliotece znajdują się tomy instruujące nas w podstawowych aspektach życia, rzeczach, które inne zwierzęta robią instynktownie lub uczą się od siebie nawzajem bez języka mówionego lub pisanego, takich jak reprodukcja, wychowywanie potomstwa, nawiązywanie przyjaźni, wydajne bieganie, skuteczne polowanie , jedzenie odpowiedniej żywności i przetrwanie "na dziko" bez telefonu komórkowego. W następnych wpisach przytłaczające dowody na pochodzenie i ewolucję człowieka oraz podstawowe koncepcje używane do interpretacji tych dowodów są przedstawione i omówione tak, jak tylko ludzie mogą to zrobić, za pomocą arbitralnego systemu symboli wydrukowanych na przetworzonej miazdze drzewnej. Czytanie o pierwszych miotaczach włóczni, prehistorycznych obrażeniach podczas polowania na mamuty lub pierwszej morskiej podróży do Australii powinno być jak czytanie o cudownym ramieniu rzucającym na twojego stryjecznego dziadka, o tym, jak twój pradziadek zginął, zataczając się przy 150-funtowej rybie, czyli o tym, jak twoja prapraprababcia wyemigrowała do Ameryki na łodzi z Irlandii. Mam nadzieję, że czytelnicy odrzucą tę ważną zasadę badań nad pochodzeniem człowieka: jesteśmy zmodyfikowanymi małpami afrykańskimi, które pomimo pozornie wielkiej różnorodności w zdobnictwie biologicznym i kulturowym, mają wspólnego afrykańskiego przodka. Wszyscy jesteśmy jednym, zróżnicowanym gatunkiem o zróżnicowanych regionalnie cechach fizycznych, które są wynikiem adaptacji środowiskowych, preferencji partnerów, migracji lub po prostu przypadku. Kultura ma znaczący wpływ na różnice, które dostrzegamy u innych ludzi. Pod koszulkami, malowaniem twarzy, kolczykami, tatuażami, irokezami i szpilkami wszyscy jesteśmy bardzo podobni. To dzięki naszemu wyjątkowemu makijażowi mamy zarówno skłonność do zapominania, jak i zdolność przyjmowania, że jesteśmy integralną częścią otaczającego nas świata przyrody.